3.15.2. Механизмы условного
торможения
Каким образом неподкрепление условного сигнала приводит к ослаблению и прекращению условной реакции? Хотя И. П. Павлов и не давал однозначного ответа на этот вопрос, но высказывал предположение о том, что условное торможение возникает и первоначально локализуется в корковых клетках «представительства условного раздражителя». Другие ученые полагали, что условное торможение правильнее всего связывать с «корковым представительством безусловного рефлекса» или со структурами, осуществляющими временную связь между условным и безусловным раздражителями.
Трудно судить о приуроченности тех
или иных центральных процессов при условном торможении к работе строго
ограниченных структур мозга. Поэтому термин «внутреннее торможение» вряд ли
имеет строгое логическое обоснование в пределах системы тормозного условного
рефлекса.
При этом следует обратить внимание
на сходство закономерностей образования положительных и тормозных условных
рефлексов. Считается, что условный рефлекс возникает в результате синтеза двух
безусловных рефлексов, один из которых является ориентировочным на действие
условного раздражителя (рис. 3.45). Предполагается, что возбуждение
безусловного рефлекса производит индукционное торможение среднего звена дуги
ориентировочного рефлекса условного раздражителя. После прекращения действия
безусловного раздражителя данное торможение сменяется послетормозным
возбуждением. Последнее, в свою очередь, вызывает отрицательное индукционное
торможение условной связи. Это торможение охранительно—восстановительного типа
предотвращает следовое перевозбуждение условной связи и утомление
соответствующих корковых клеток. При отмене подкрепления нарушается
|
|
|
Рис. 3.45 Дуга
условного рефлекса с двусторонней связью А, Б — корковые
представительства мигательного и пищевого рефлексов; В, Г — подкорковые представительства мигательного и пищевого
рефлексов: 1,1' — афферентные
нейроны, 2, 2' — вставочные
нейроны, 3, 3' — эфферентные нейроны, 4,
5, 4', 5' — синаптические контакты коллатералей афферентных нейронов, 6 — синаптический контакт между
вставочным нейроном обратной условной связи и афферентным нейроном
сигнального раздражителя, 6' — синаптический
контакт между вставочным нейроном Прямой условной связи и афферентным нейроном
подкрепляющего раздражителя, I ,II — прямая и обратная условные связи. |

сопряженная индукция и элементы
условной связи испытывают длительное следовое возбуждение.
Согласно биологической гипотезе,
в основе механизма условного торможения лежит конкурентная борьба, столкновение
различных возбуждений. Более сильное возбуждение индукционно тормозит
выработанный пищевой рефлекс. Если при безусловном торможении такое возбуждение
возникает при ориентировочном или оборонительном рефлексе, то при условном
торможении — при биологически отрицательной реакции.
На рис. 3.46 приведена схема образования
угасательного торможения. Общий механизм тормозящего эффекта определяет единую
природу коркового торможения, в основе которого может лежать пессимальное, а
иногда и электротоническое торможение.
Считается, что при выработке
условного торможения формируется самостоятельный тормозный условный рефлекс,
независимый от положительного.
Приведенные гипотезы построены в
значительной мере на косвенных данных, а поэтому носят умозрительный характер,
нисколько не снижающий, впрочем, их оригинальной ценности.
Клеточный анализ условного
торможения показал, что в формировании тормозной реакции участвуют все звенья
положительного условного рефлекса, но они работают в другом режиме, меняются
параметры их деятельности. Отсутствие внешней реакции на отрицательный
(неподкрепляемый) стимул не связано с локальным или системным торможением
структур головного мозга. В ответ на тормозное раздражение возникает такая же
многообразная активность нейронов, как на положительное раздражение.
Некоторые исследователи видят
проявления условного торможения в снижении или прекращении импульсной
активности клетки при продолжительном действии положительного условного сигнала
(торможение запаздывания). Обнаружено, что при угасательном торможении
амплитуда ВПСП постепенно снижается без заметного перехода в гиперполяризацию
клеточной мембраны. Появилась возможность объяснить ослабление возбуждающих
реакций уменьшением выброса медиатора без активного торможения самого нейрона.
Подобные наблюдения, а также факты постепенного угнетения клеточных реакций при
угашении могут быть связаны с понижением возбудимости корковых
|
|
|
Рис. 3.46 Образование угасательного торможения А — условный пищевой рефлекс; Б — стадия частичного
торможения пищевой реакции вследствие появления биологически отрицательной
реакции; В — стадия полного торможения пищевой реакции: / — анализатор, 2 — корковое представительство
пищевой реакции, 3 — корковое представительство биологически отрицательной
реакции, 4— путь для
побочного торможения |
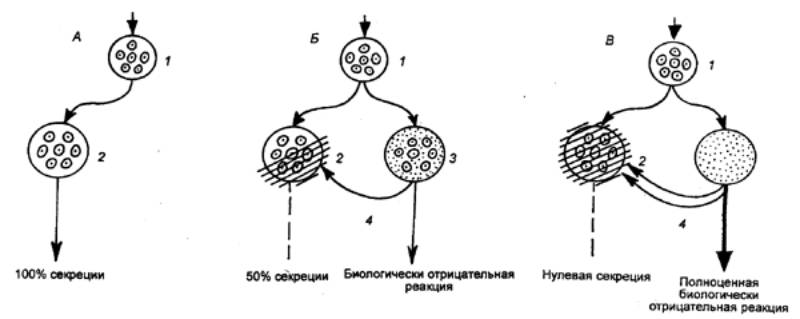
нейронов. Вероятно, при развитии
угасательного торможения имеет место размыкание временной связи.
Положительные и отрицательные
условные рефлексы опосредуются разными синаптическими входами активирующего и
тормозного действия. В зависимости от того, какие интернейроны включены в
полисинаптические пути условной связи, дифференцировочное торможение будет
проявляться или как угнетение активирующих условных реакций клетки, или как
растормаживание клеточных реакций с тормозной условной реакцией.
Самостоятельная группа клеток на неподкрепляемое раздражение усиливала
импульсацию, а на подкрепляемый положительный сигнал не реагировала вовсе.
При выполнении инструментальных
условных рефлексов на комплексный сигнал вырабатывали условное торможение
реакций при изолированном предъявлении компонентов (рис. 3.47). Установлено,
что паттерн ответа этих нейронов на положительный сигнал (свет + звук)
существенно отличался от такового при дифференцировочных сигналах (свет, звук).
Эти паттерны ответов обусловлены двигательной программой, которая реализуется
по данному сигналу. Если угасательное торможение сопровождается распадом
паттерна нейронных ответов и/или их исчезновением, то дифференцировочное
торможение, тренируемое вместе с положительным рефлексом, стабильно
поддерживает новый паттерн нейронного ответа, отражающий центральную программу
поведенческой реакции.
При разных видах условного
торможения существует три типа перестроек импульсации: 1) инверсия
паттерна активности нейрона, имеющая место при положительном сигнале, в
противоположный по знаку при предъявлении отрицательного сигнала; 2)
рекомбинация паттерна ответа при предъявлении неподкрепляемого раздражителя в
сравнении с эффектом положительного сигнала; 3) исчезновение реакции нейронов
при предъявлении дифференцировочного раздражителя. Эти перестройки именуются
как центральный нейронный эквивалент дифференцировочного торможения.
Условное торможение является таким
же активным системным процессом, как и условное возбуждение. По своей
организации, за исключением конечного эффекторного звена, оба процесса подобны.
Вероятно, мозаика постсинаптических
процессов, происходящих на корковом уровне, отражает перестройки функциональной
активности соответствующих центральных программ, эффекторное выражение которых
осуществляется на соответствующих
|
|
|
Рис. 3.47 Перистимульная гистограмма активности нейрона сенсомоторной
коры кошки при положительных и отрицательных условных рефлексах А — положительные условные
рефлексы на комплексный сигнал из одновременных светового и звукового
компонентов; Б, В — отрицательные условные рефлексы
на изолированные предъявления светового и звукового компонетов. По оси
ординат — число импульсов, каждый столбик соответствует периоду в 50 мс, по
оси абсцисс — время в секундах, каждая гистограмма построена на основе 10
реализации; под гистограммой: отклонение вверх — условный сигнал, вниз —
подача подкрепления, горизонтальные отрезки — проявление двигательной реакции
передней конечности. |

|
|
|
Рис. 3.48
Временная организация памяти. (Объяснение см. в тексте). |
