Спинной мозг (рис. 3.5.) является филогенетически
наиболее древним отделом ЦНС. В примитивной форме у ланцетника он
представляет собой трубку, идущую вдоль всего тела. От спинного мозга отходят
вентральные (передние, или двигательные) и дорсальные (задние, или
чувствительные) корешки (рис. 3.6).
У ланцетника спинномозговых узлов (спинальных ганглиев) еще
нет, чувствительные клетки рассеяны по ходу нервов или лежат в дорсальных
отделах спинного мозга. У круглоротых уже намечается разделение спинного мозга
на серое вещество, содержащее клетки и составляющее центральную часть, и
окружающее его белое вещество из продольно идущих безмякотных волокон.
Появляются и спинномозговые узлы. Они лежат на дорсальных корешках и состоят из
биполярных чувствительных нейронов.
Более сложно устроен спинной мозг круглоротых. В связи с тем
что у них происходит миелинизация нервных волокон, серое и белое вещество
приобретают четкие границы. В сером веществе возникают вентральные и дорсальные
рога, в белом веществе — вентральные и латеральные канатики. Вентральный и
дорсальный корешки сливаются и дают начало смешанному нерву. Таким образом, спинной
мозг круглоротых является прототипом спинного мозга высших позвоночных.
С появлением конечностей у амфибий возникают шейное и
поясничное утолщения спинного мозга, в передних рогах серого вещества выделяются
медиальная и латеральная группы клеток, образуются нисходящие и восходящие
пути.
|
|
|
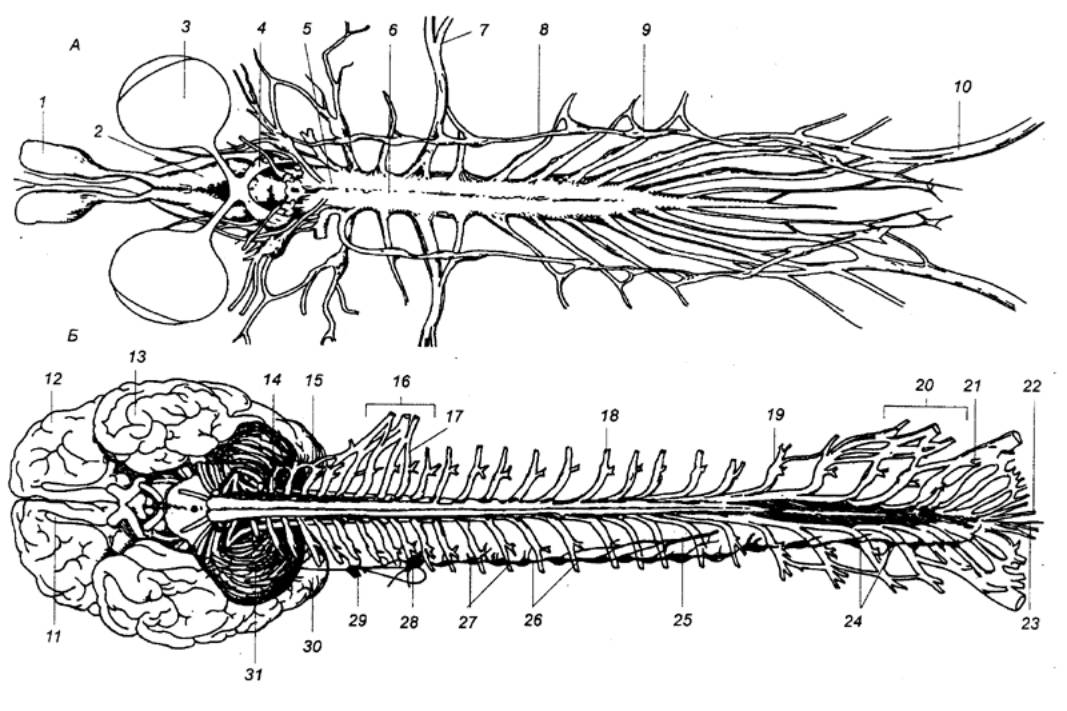
Рис. 3.5 Спинной
мозг лягушки (А) и человека (Б) с вентральной стороны 1 — носовой мешок, 2 — головной мозг, 3 — глаз, 4 — зрительный тракт, 5 —
продолговатый мозг, 6 —
спинной мозг, 7 — плечевой нерв, 8 — вегетативный ствол, 9 — спинальный ганглий, 10 — седалищный нерв, 11 — обонятельная луковица, 12 — лобная доля, 13 — височная доля, 14 — 1—й шейный нерв, 15 — мозжечок, 16 — плечевое сплетение, 17 — 1—й грудной нерв, 18 —спинальный ганглий, 19 — 1—й поясничный нерв, 20 — поясничное сплетение, 21 — 1—й крестцовый нерв, 22— копчиковый нерв, 23—концевая нить, 24 — соединительные ветви, 25 — чревный нерв, 26 — симпатический ствол, 27 — симпатический ганглий, 28 — нижний шейный симпатический
ганглий, 29 — средний шейный
симпатический ганглий, 30 —
затылочная доля, 31 — верхний
шейный симпатический ганглий. |
|
|
|
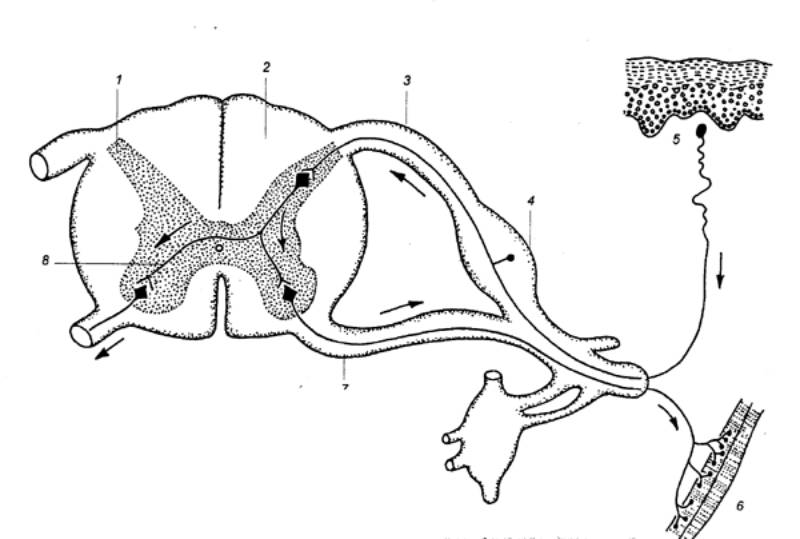
Рис. 3.6
Спинной мозг и его взаимосвязь с периферическими нервными волокнами— чувствительным,
двигательным. 1 — серое
вещество, 2 — белое вещество,
3 — дорсальный (задний) корешок, 4 —
спинальный ганглий, 5 — рецептор, 6
— эффектор, 7 —
вентральный (передний) корешок, 8 —
интернейрон. Стрелками показано направление распространения возбуждения. |
Дальнейшая дифференцировка
клеточно—волоконных структур происходит у рептилий и птиц. У них получают
развитие восходящие пути, лежащие в центральном и боковом канатиках, сформированы
дорсальный чувствительный путь и нисходящие связи. Происходит также
дифференциация ассоциативных клеток, возникновение межсегментарных
комиссуральных связей. У птиц хорошо развиты связи спинного мозга с
вестибулярным аппаратом и мозжечком.
Еще большая дифференцированность серого и белого вещества
наблюдается у млекопитающих. В дорсальных рогах появляются студенистое вещество
и грудное ядро, в вентральных — отчетливо выделяются клеточные группы,
возрастает число волокон в дорсальных и вентральных канатиках, появляется новый
восходящий путь от шейных сегментов к оливам.
В организации спинного мозга млекопитающих существуют
особенности, связанные с видовой принадлежностью животных. Спинной мозг у них
имеет разную длину, разное количество сегментов, неодинаковую выраженность
утолщений и пр. Все это зависит от числа позвонков, наличия хвоста, функции
конечностей.
При изучении деятельности спинного мозга необходимо
учитывать, что тело позвоночных животных и человека может быть разделено на пояса,
или сегменты. Сегменты, получающие чувствительные волокна от одной
отдельной пары дорсальных корешков, образуют метамер. Кожная область, в
которой распределяются эти чувствительные волокна, называется дерматомом
(рис. 3.7).
|
|
|
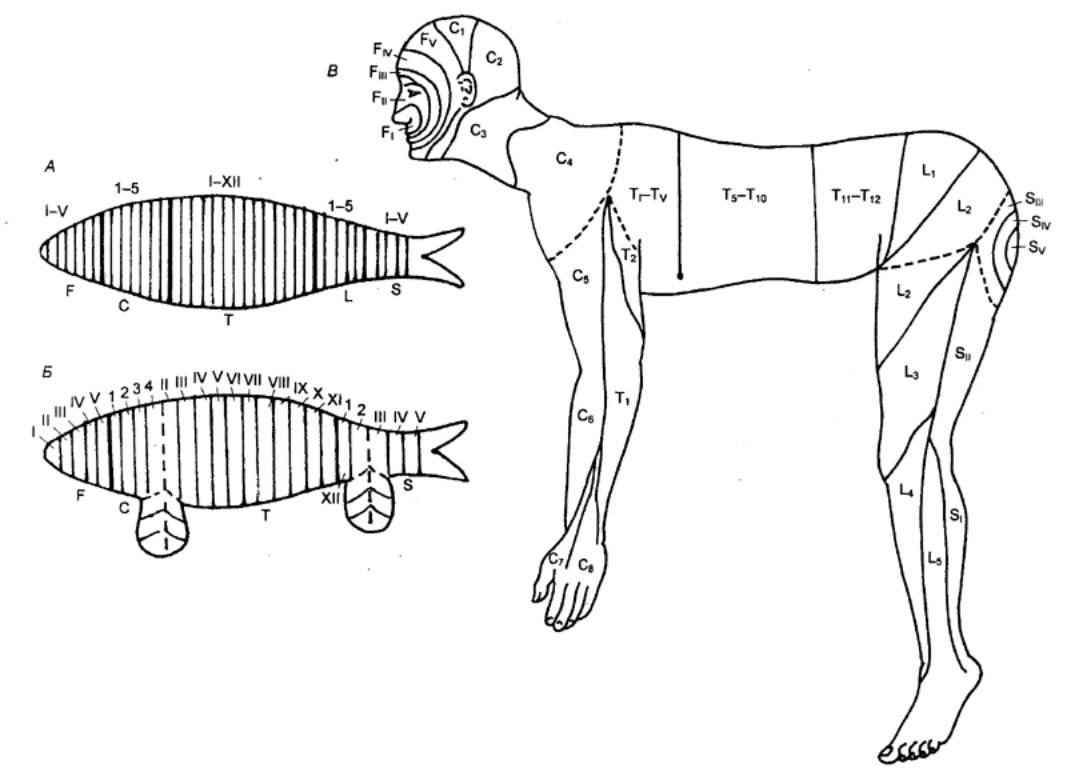
Рис. 3.7 Распределение дерматомов (областей чувствительной
иннервации) на поверхности тела Буквы соответствуют отделам спинного мозга: С — шейному, Т
— грудному, L — поясничному и S — крестцовому; цифрами показаны номера сегментов.
От каждого участка кожи, обозначенного одинаковыми символами, чувствительные
волокна отходят в один сегмент спинного мозга, например, чувствительность
кожи в области СЗ шеи обеспечивается 3—м шейным сегментом, область L5 на голени — 5—м поясничным
сегментом. |

|
|
|
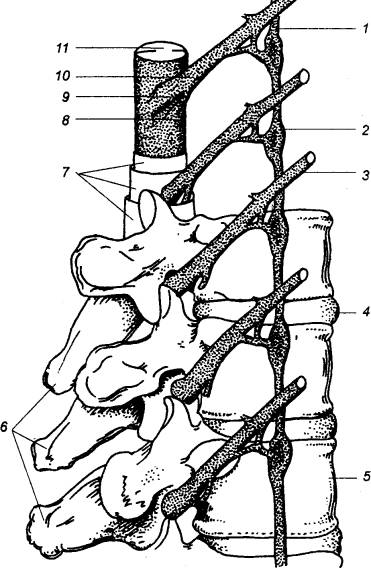
Рис. 3.8 Позвоночный столб, спинной мозг и пограничный
симпатический ствол 1 — симпатический ствол, 2 — симпатический ганглий, 3 —
спинномозговой нерв, 4 —
межпозвоночный диск, 5 —
позвонок, 6 — остистные
отростки позвонков, 7 — соединительнотканные оболочки, 8 — вентральный корешок, 9 — спинальный ганглий, 10 — дорсальный корешок, 11 — спинной мозг. |
В процессе эволюции тело позвоночных
изменило строение, в итоге возникли существенные отступления от идеальной метамерии.
Например, у ланцетника метамерия делит длину тела на неодинаковые отрезки.
Перестройку метамеров вызвало появление плавников у рыб. Метамерность особенно
усложнилась в связи с развитием конечностей и приобрела сложную форму для рук и
ног человека (рис. 3.7, 3.9).
Спинной мозг человека состоит из
следующих сегментов (обозначаются латинскими буквами): 8 шейных —C(I—8) 12 грудных — Т(1—12), 5 поясничных — L(1—5), 5 крестцовых —
S(1—5), 3 копчиковых — Со(1—3).
Общее количество сегментов соответствует числу метамеров тела, однако каждый
метамер получает иннервацию от двух—трех лежащих рядом сегментов. В сегментах
спинного мозга заканчиваются отростки подавляющего большинства чувствительных
нейронов тела, вступающие в составе дорсальных корешков. В спинном мозгу также
начинаются почти все эфферентные нервы организма: двигательные (за исключением
иннервирующих мышцы головы), все симпатические и часть парасимпатических. Они
следуют в составе дорсальных и вентральных корешков (рис. 3.8).
Если у лягушки перерезать справа
дорсальные, а слева вентральные корешки пояснично—крестцовых сегментов, то
правая лапка полностью теряет чувствительность, но может совершать движения.
Левая, наоборот, сохранит чувствительность, но окажется не способной к
двигательным реакциям. Это явление было открыто независимо друг от друга Ч.
Беллом (1811) и Ф. Мажанди (1822) и получило название закона Белла—Мажанди.
Характерной особенностью является
соотношение волокон, входящих через дорсальные и выходящих через вентральные
корешки. У кошки, например,
|
|
|
Рис.3.9 Схема
метамерности тела в процессе эволюции.
А— правильная
последовательность расположения метамеров (теоретически возможный случаи для
позвоночных); Б — нарушение правильной
метамерности тела рыб в связи с образованием плавников; В — то же у человека в связи с
образованием конечностей; F —
сегменты головы; С — сегменты шеи; Т — грудные сегменты; L — поясничные сегменты; S — крестцовые сегменты. |
дорсальный корешок поясничного
сегмента содержит около 12 000 волокон, тогда как вентральный — 6000.
Следовательно, один и тот же двигательный нейрон является общим конечным путем
для импульсов, поступивших от разных рецепторов и конкурирующих за общий конечный
путь. Несмотря на такую организацию, степень интегративной деятельности
спинного мозга ограничена по сравнению с другими вышележащими отделами ЦНС.
Классическими методами изучения функций спинного мозга
являются перерезки или разрушения его структур с последующей оценкой нарушения
функций. Большое развитие получило применение электрофизиологических методов,
включающих регистрацию суммарных электрических процессов, вне— и
внутриклеточные методы регистрации активности отдельных клеток.
3.3.1. Нейронные структуры и их свойства
Тела чувствительных клеток вынесены за пределы спинного
мозга. Часть из них располагается в спинномозговых узлах (спинальных ганглиях)
(рис. 3.10, 3.11). Это тела соматических афферентов, иннервирующих в основном
скелетные мышцы. Другие находятся в экстра— и интрамуральных ганглиях
автономной нервной системы и обеспечивают чувствительность только внутренних
органов.
Чувствительные клетки имеют один отросток, который вскоре
после отхождения от тела клетки делится на две ветви (псевдоуниполярный
нейрон). Одна из них проводит возбуждение от рецепторов к телу клетки, другая —
от тела
|
|
|
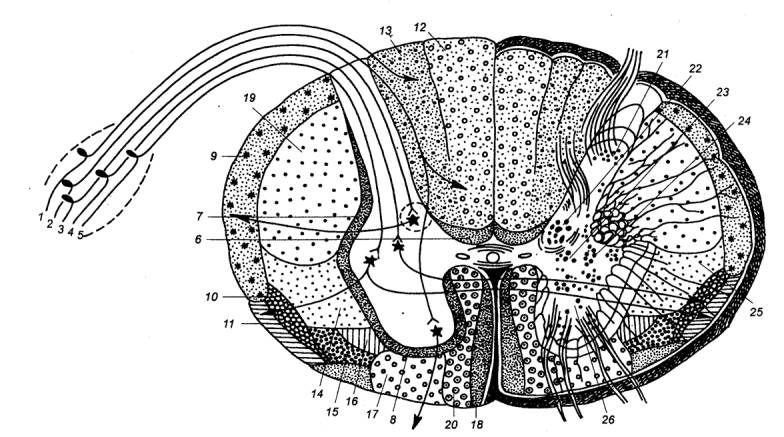
Рис.3.10
Схема поперечного разреза спинного мозга и спинального ганглия.1,2 — рефлекторные пути сознательных
проприоцептивных ощущений и осязания, 3,4
— рефлекторные пути проприоцептивных импульсов, 5 — рефлекторные пути
температурной и болевой чувствительности, 6 — задний собственный пучок, 7 — боковой собственный пучок, 8 — передний собственный пучок, 9 — задний спинно—мозжечковый путь, 10 — передний спинно—мозжечковый путь, 11 — спинно—таламический путь, 12 — тонкий пучок, 13
— клиновидный пучок, 14 —
красно—ядерно—спинномозговой путь, 15
— таламо—спинномозговой путь, 16
— преддверно—спинномозговой путь, 17 — ретикулярно—спинномозговой путь, 18 — покрышечно—спинномозговой путь, 19 — латеральный корково—спинномозговой (пирамидный) путь, 20 — передний
корково—спинномозговой (пирамидный) путь, 21 — собственное ядро заднего рога, 22 — грудной столб (ядро), 23, 24 — промежуточное (серое) вещество, 25 — боковой промежуточный столб, 26 — ядра переднего рога. |
|
|
|
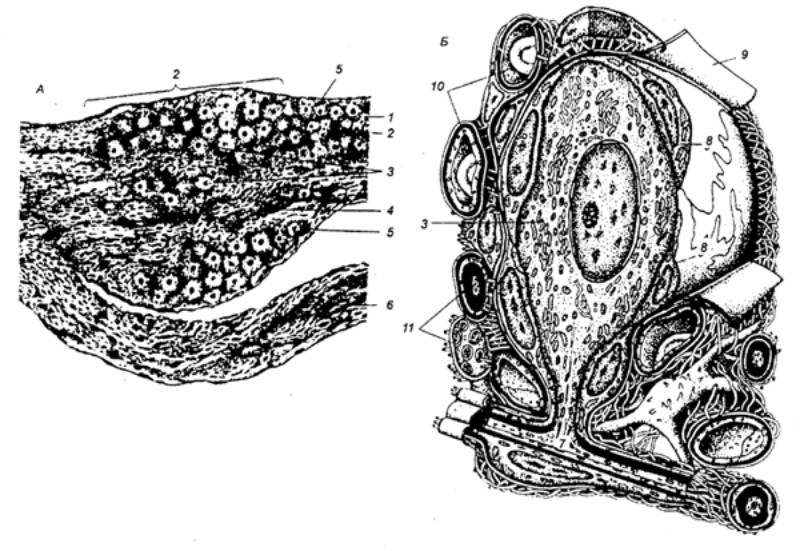
Рис. 3.11
Строение спинального ганглия.А —
продольный разрез ганглия и корешков; Б
— ультраструктурная организация псевдоуниполярного нейропита и его
микроокружения; 1 — дорсальный
корешок; 2 — ганглий; 3 —
псевдоуниполярные нейроциты; 4 —
нервные волокна; 5 — соединительнотканная капсула; 6 — вентральный корешок; 7 — Т—образное разветвление
отростка; 8 — мантийные
глиоциты; 9 — бааальная
мембрана; 10 — гемокапилляры; 11 —
миелиновые и безмиелиновые нервные волокна. |
|
|
|
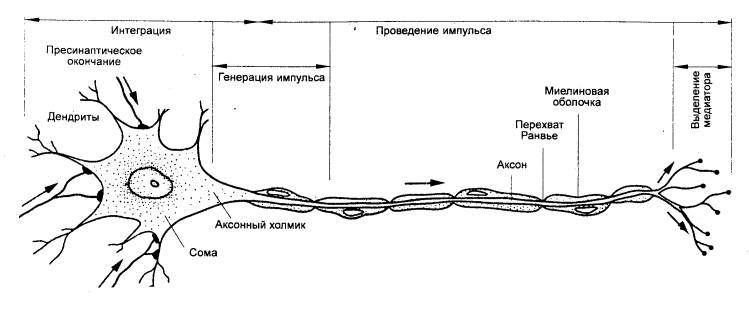
Рис. 3.12 Мотонейрон
спинного мозга позвоночных. Указаны функции разных его частей |
нервной клетки к нейронам спинного или
головного мозга. Распространение возбуждения из одной ветви в другую может
происходить без участия тела клетки.
Нервные волокна чувствительных
клеток по скорости проведения возбуждения и диаметру классифицируют на А—, В—
и С—группы. Толстые миелинизированные А—волокна с диаметром от 3 до 22
мкм и скоростью проведения возбуждения от 5 до 120 м/с разбиваются еще на
подгруппы: а — волокна от мышечных рецепторов, Р — от тактильных рецепторов и
барорецепторов, 8 — от терморецепторов, механорецепторов, болевых рецепторов. К
волокнам группы В относят миелиновые отростки средней толщины со
скоростью проведения возбуждения 3—14 м/с. По ним в основном передается
ощущение боли. К афферентным волокнам типа С относится большинство
безмиелиновых волокон толщиной не более 2 мкм и скоростью проведения 0,5—2 м/с.
Это волокна от болевых, хемо— и некоторых механорецепторов.
Сам спинной мозг в целом содержит, например, у человека
примерно 13 млн. нейронов. Из их общего числа только около 3% составляют
эфферентные, двигательные, или моторные, нейроны (рис. 3.12), а остальные 97%
приходятся на долю вставочных, или интернейронов. Двигательные нейроны являются
выходными клетками спинного мозга. Среди них различают α— и
γ—мотонейроны, a также преганглионарные нейроны автономной нервной системы.
α—Мотонейроны осуществляют передачу скелетным мышечным волокнам сигналов,
выработанных в спинном мозгу. Аксоны каждого мотонейрона многократно делятся,
и, таким образом, каждый из них охватывает своими терминалями до сотни мышечных
волокон, образуя в совокупности с ними двигательную единицу. В свою
очередь, несколько мотонейронов, иннервирующих ,одну и ту же мышцу, образуют мотонейронный
пул. В его состав могут входить мотонейроны нескольких соседних сегментов.
В связи с тем, что возбудимость / мотонейронов пула неодинакова, при слабых
раздражениях возбуждается только часть из них. Это влечет за собой
соответственно сокращение лишь части мышечных волокон. Другие моторные единицы,
для которых это раздражение является подпороговым, тоже реагируют, хотя их
реакция выражается лишь в деполяризации мембраны и повышении возбудимости. С
усилением раздражения они еще больше вовлекаются в реакцию, и, таким образом,
все двигательные единицы пула участвуют в рефлекторном ответе.
Максимальная частота воспроизведения ПД в α—мотонейроне
не превышает 200—300 имп/с. Вслед за ПД, амплитуда которого составляет 80—100
мВ, возникает следовая гиперполяризация длительностью от 50 до 150 мс.
По частоте импульсации и выраженности следовой гиперполяризации мотонейроны
разделяют на две группы: фазические и тонические. Особенности их возбуждения
коррелируют с функциональными свойствами иннервируемых мышц. Фазическими
мотонейронами иннервируются более быстрые, «белые» мышцы, тоническими —
более медленные, «красные» (см. разд. 1.2.1).
В организации функции α—мотонейронов важным звеном
является наличие системы отрицательной обратной связи, образованной
аксонными коллатералями и специальными тормозными вставочными нейронами — клетками
Реншоу. Своими возвратными тормозными влияниями они могут охватывать большие
группы мотонейронов, обеспечивая, таким образом, интеграцию процессов
возбуждения и торможения.
γ— Мотонейроны, иннервируют интрафузальные
(внутриверетенные) мышечные волокна. Они разряжаются с более низкой частотой, и
следовая гиперполяризация у них выражена слабее, чем у α—мотонейронов. Их
функциональное значение сводится к сокращению интрафузальных мышечных волокон,
не приводящему, однако, к появлению двигательного ответа. Возбуждение этих
волокон сопровождается изменением чувствительности их рецепторов к сокращению
или расслаблению экстрафузальных мышечных волокон.
Нейроны автономной нервной системы составляют особую группу клеток.
Тела симпатических нейронов, аксоны которых являются преганглионарными
волокнами, располагаются в боковом промежуточном столбе спинного мозга (см.
разд. 5.1.2). По своим свойствам они относятся к группе В—волокон. Характерной
особенностью их функционирования является низкая частота свойственной им
постоянной тонической импульсной активности. Одни из этих волокон участвуют в
поддержании сосудистого тонуса, другие — обеспечивают регуляцию висцеральных
эффекторных структур (гладкой мускулатуры пищеварительной системы, железистых
клеток).
Тела парасимпатических нейронов образуют крестцовые
парасимпатические ядра. Они располагаются в сером веществе крестцовых сегментов
спинного мозга. Для многих из них характерна фоновая импульсная активность,
частота которой возрастает по мере повышения давления в мочевом пузыре. При
раздражении висцеральных тазовых афферентных волокон в эфферентных клетках
регистрируется вызванный разряд, характеризующийся чрезвычайно большим
латентным периодом.
К вставочным, или интернейронам, спинного
мозга относятся нервные клетки, аксоны которых не выходят за его пределы. В
зависимости от хода отростков различают собственно спинальные и проекционные. Спинальные
интернейроны ветвятся в пределах нескольких смежных сегментов, образуя
внутрисегментарные и межсегментарные связи. Наряду с ними существуют
интернейроны, аксоны которых проходят через несколько сегментов или даже из
одного отдела спинного мозга в другой. Их аксоны образуют собственные пучки
спинного мозга.
К проекционным интернейронам
относятся клетки, длинные аксоны которых формируют восходящие пути спинного
мозга. На каждом интернейроне расположено в среднем около 500 синапсов.
Синаптические влияния в них опосредуются через ВПСП и ТПСП, суммация которых и
достижение критического уровня приводят к возникновению распространяющегося ПД.
Дорсальную часть дорсального рога спинного мозга занимает
характерное скопление нервных клеток, образующих густое сплетение. Эта область
получила название студенистого (роландова) вещества. У
позвоночных животных и человека послойное рассмотрение клеточной архитектуры
серого вещества спинного мозга показало существование последовательно
расположенных слоев, или пластин, состоящих из однотипных клеток.
Согласно Рекседу (Rexed), все серое вещество разделяется на 10 таких пластин (рис.
3.13).
|
|
|
Рис. 3.13 Послойная топография
серого вещества спинного мозга по Рекседу |
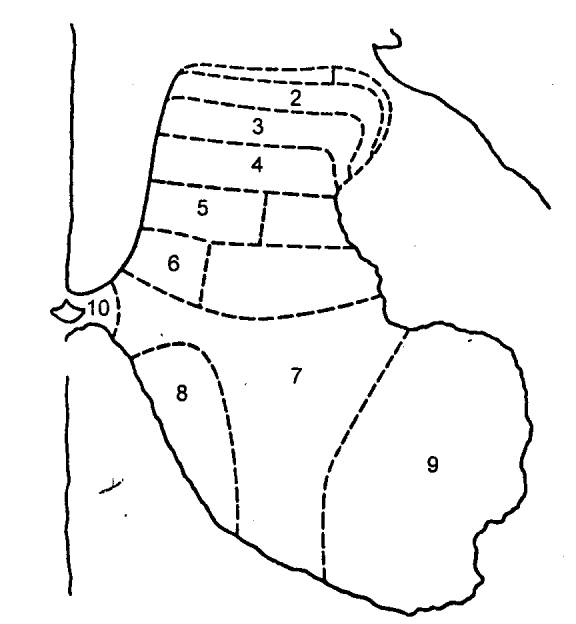
Из них первая — дорсальная, содержит преимущественно
нейроны, аксоны которых дают начало спинно—таламическому пути. На самих нейронах
оканчиваются первичных афференты и аксоны проприоспинальных нейронов. Вторая
и третья пластины образуют названное выше студенистое вещество. Здесь
выделяются
два вида нейронов с многочисленным
дендритным ветвлением — мелкие и крупные. Аксоны этих клеток в большинстве
оканчиваются здесь же в студенистом веществе, хотя некоторые из них
проецируются на дорсолатеральный собственный пучок спинного мозга. Четвертая
пластина локализуется в центре дорсального рога. Составляющие ее клетки
получают синаптический приток от нейронов студенистого тела и первичных
чувствительных нейронов, аксоны же проецируются в таламус и боковое шейное
ядро. Пятая и шестая пластины представлены конгломератом
разнообразных вставочных нейронов, получающих синаптический приток от волокон
дорсальных корешков, а также и от нисходящих путей — в основном
кортиково—спинномозгового и
красноядерно—спинномозгового путей. Седьмая и восьмая
пластины являются местом положения проприоспинальных вставочных нейронов с
длинными аксонами. На этих интернейронах оканчиваются аксоны проприорецепторов
и проприоспинальных нейронов, а также волокна преддверно—спинномозгового и
ретикулярно—спинномозгового путей. Девятая пластина является местом
сосредоточения α— и γ—мотонейронов. Этой зоны достигают пресинаптические
структуры первичных афферентов от мышечных рецепторов растяжения. Сюда приходят
также окончания волокон нисходящих трактов, аксоны возбуждающих и тормозящих
интернейронов. Наконец, десятая пластина. Она окружает спинномозговой
канал и характеризуется наряду с присутствием нервных клеток еще и наличием
значительного количества глиальных и комиссуральных волокон.
3.3.2. Рефлекторная функция спинного мозга
Важную роль в развитии представлений о рефлекторной
деятельности спинного мозга сыграли открытия и обобщения английского физиолога
Ч. Шеррингтона.
Объем функций, осуществляемых спинным мозгом, чрезвычайно
велик. В нем находятся центры: всех двигательных рефлексов (за исключением мышц
головы); всех рефлексов мочеполовой системы и прямой кишки; рефлексов,
обеспечивающих терморегуляцию; регулирующих метаболизм тканей; большинства
сосудистых рефлексов; сокращения диафрагмы и др. В естественных условиях эти
рефлексы всегда испытывают влияние высших отделов головного мозга.
Степень проявления рефлексов зависит от того, сохраняются ли
связи структур спинного мозга со структурами головного мозга. После децеребрации
(удаления головного мозга) или спинализации (отделения спинного мозга от
головного посредством перерезки) исчезают многие сложные формы активности,
создаваемые спинным мозгом. При этом определенное значение принадлежит уровню
организации подопытного животного. Например, спинальная лягушка, представитель
низших позвоночных, может сидеть и вырываться, когда ее схватывают. Спинальная
же собака сама не может ни стоять, ни ходить. Это объясняется тем, что
разобщение спинного мозга и вышележащих структур нарушает рефлекторные дуги,
ответственные за осуществление определенных реакций. При этом, в частности,
исчезают периодические разряды дыхательных мышц, обеспечивающие дыхательные
движения, пропадают тонические разряды симпатических нейронов, поддерживающих
сосудистый тонус и, соответственно, артериальное давление.
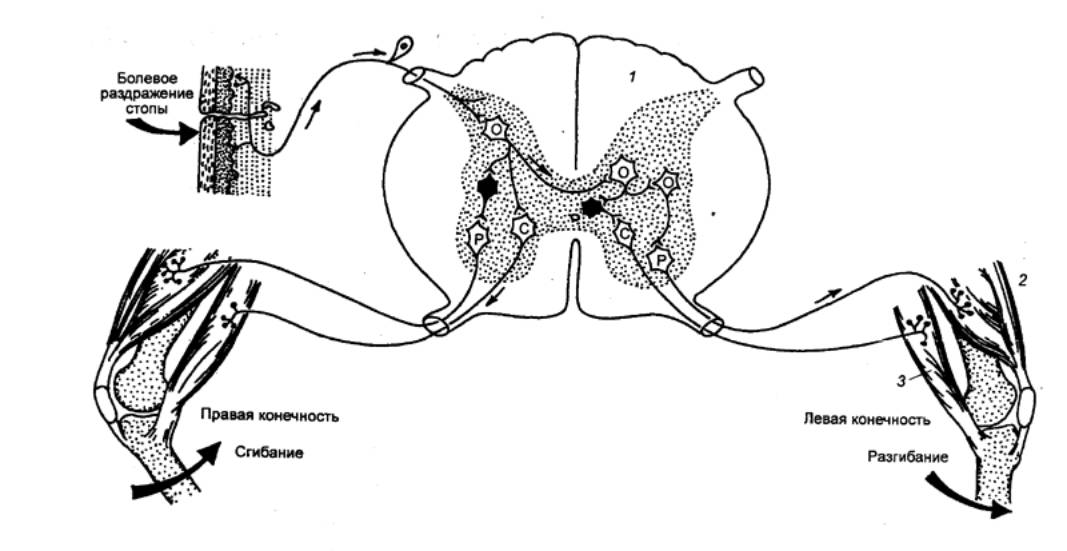
Как правило, рефлексы спинальных животных являются
координированными. Возбуждение у них каждой группы рецепторов сопровождается
своим специфическим ответом. Например, механическое раздражение кожи стопы у
лягушки вызывает сгибание раздражаемой конечности и разгибание другой.
Раздражение рецепторов мочевого пузыря и прямой кишки сопровождается рефлекторным
сокращением их мускулатуры.
В связи с отсутствием тонических влияний структур головного
мозга изменяется функциональное состояние и самих спинномозговых нейронных
систем. К числу таких нарушений относится прекращение сложных локомоторных
актов типа шагания. Характерно, что исчезновение этих актов после спинализации
удается восстановить введением животному веществ, способствующих выделению
медиаторов синаптическими окончаниями перерезанных путей.
|
|
|
Рис. 3.14 Внутрисегментарные связи сенсорного волокна от
болевого рецептора кожи стопы Р —
мотонейроны разгибателей; С — мотонейроны сгибателей. |
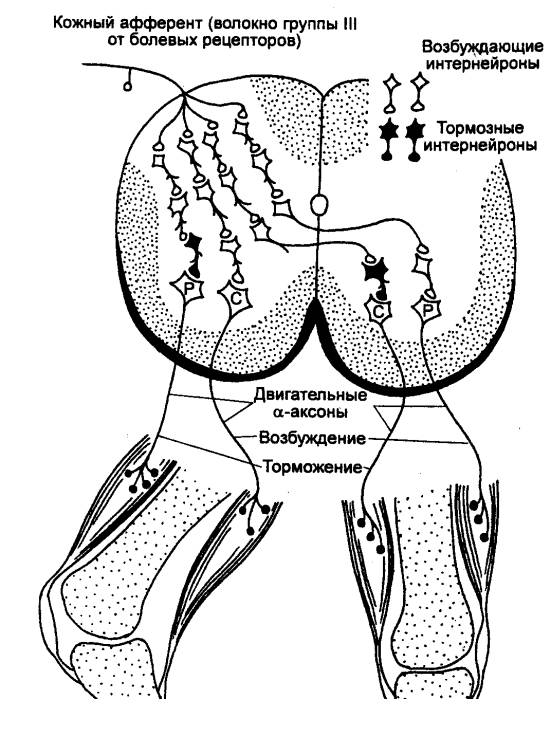
В зависимости от числа нейронов, участвующих в проведении возбуждения,
рефлекторные дуги спинного мозга делятся на моносинаптические и
полисинаптические.
Моносинаптическая дуга состоит из чувствительного нейрона
с рецепторами мышечных веретен и эффекторного нейрона, оканчивающегося на
мышечных волокнах. Классическим примером моносинаптической дуги может служить
рефлекторная дуга коленного рефлекса, в которой возбуждение от рецептора до
эффектора проходит всего за 0,5—1,0 мс, т. е. время, необходимое для
прохождения возбуждения всего лишь через один синапс.
В полисинаптической дуге на
пути возбуждения от рецептора к эффектору помимо чувствительного и эффекторного
нейрона находятся еще вставочные нейроны (рис. 3.14). Таким образом,
возбуждение в этой дуге проходит не через один, а через несколько синапсов,
определяющих время латентного периода ответа и суммарной синаптической
задержки. При осуществлении рефлекторных реакций
|
|
|
Рис. 3.15 Рефлексы, спинного мозга А — коленный; Б — ахиллов; В — подошвенный; Г — сгибательно—локтевой;
Д — разгибательно—локтевой; Е — брюшной. |
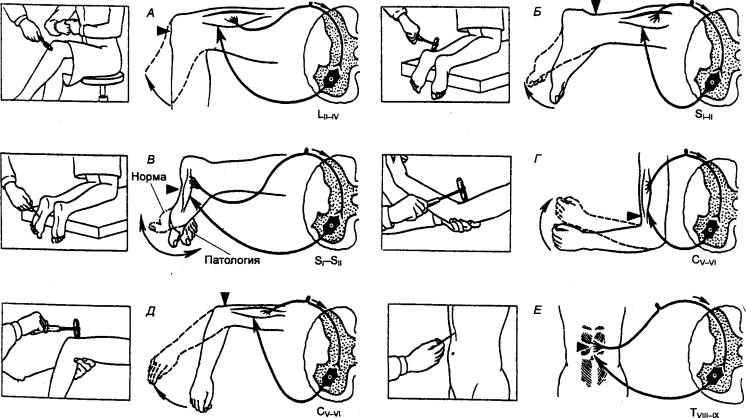
Таблица 3.1
Соматические спинномозговые рефлексы человека
|
Название
рефлекса |
Применяемое
раздражение |
Характер
рефлекторной реакции |
Локализация
нейронов, участвующих в рефлексе |
|
Сухожильные
проприорецептивные рефлексы: |
|
|
|
|
сгибательно—локтевой
(бицепс—рефлекс) |
Удар
молоточком по сухожилию двуглавой мышцы плеча (рука слегка согнута в локте) |
Сокращение
двуглавой мышцы плеча и сгибание руки |
V—VI шейные сегменты спинного мозга |
|
коленный |
Удар
молоточком по сухожилию четырехглавой мышцы бедра ниже надколенника |
Сокращение
четырехглавой мышцы бедра и разгибание голени |
II—IV поясничные сегменты |
|
ахиллов |
Удар по
ахиллову сухожилию |
Подошвенное
сгибание стопы |
1—11
крестцовые сегменты |
|
Брюшные
рефлексы: |
Штриховое
раздражение кожи: |
Сокращение
соответствующих участков брюшной мускулатуры |
|
|
верхний |
параллельно
нижним ребрам |
|
VIII—IX грудные сегменты |
|
средний |
на уровне
пупка (горизонтально) |
|
IX—XII грудные сегменты |
|
нижний |
параллельно
паховой складке |
|
I—II поясничные сегменты |
|
Кремастерный
яичковый рефлекс |
Штриховое
раздражение внутренней поверхности бедра |
Сокращение
мышцы, поднимающей яичко |
I—II поясничные сегменты |
|
Анальный
рефлекс |
Штрих
или укол вблизи заднего прохода |
Сокращение
наружного сфинктера прямой кишки |
IV—V крестцовые сегменты |
|
Подошвенный
рефлекс |
Слабое
штриховое раздражение подошвы |
Сгибание
пальцев и стопы |
I—II крестцовые сегменты |
|
Сильное
раздражение подошвы |
Разгибание
пальцев и сгибание ноги |
в моно— и полисинаптических дугах
вовлекаются также координирующие рефлекс многочисленные интернейроны. Подобный анализ
периферических воздействий, а также надсегментарный контроль за рефлекторной
деятельностью спинного мозга происходит с помощью длинных многоэтажных
рефлекторных дуг. Их центры локализованы в подкорковых областях и коре
головного мозга.
К числу рефлексов спинного мозга (рис. 3.15, табл.
3.1) относятся защитные рефлексы, рефлексы на растяжение, мышц—антагонистов,
висцеромоторные, вегетативные рефлексы. Эта классификация весьма условна, и
весь ее смысл в том, что она указывает на многообразие рефлекторных ответов.
Даже у спинального животного трудно встретить рефлексы, которые относились бы
только к одной из названных групп.
Раздражение в виде укола кожи стопы вызывает у лягушки защитный
рефлекс — либо отдергивание лапки при легком уколе, либо вовлечение в
реакцию другой лапки и отодвигание от источника при более сильном воздействии,
наконец, убегание животного при значительном болевом раздражении, когда в
возбуждение вовлекаются многие структуры нервной системы.
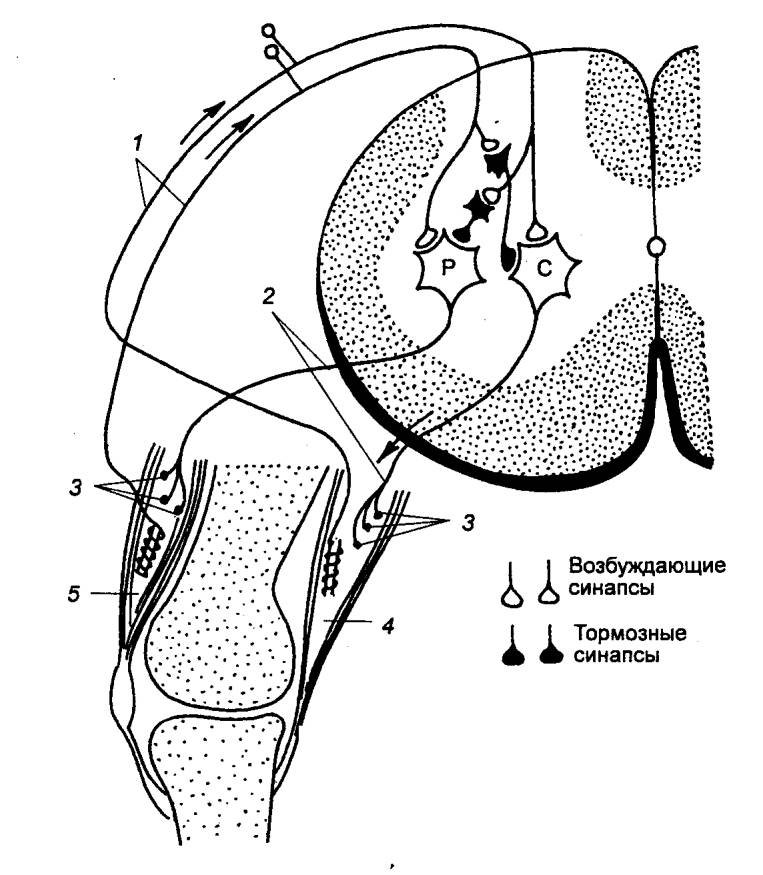
Рефлексы растяжения проявляются укорочением мышцы в
ответ на ее растяжение (рис. 3.16). Основными рецепторами в этом случае служат
нервно—мышечные веретена, афферентным звеном — чувствительные волокна
соматических нервов и дорсальных корешков спинного мозга. Эти рефлекторные дуги
чаще всего замыкаются в спинном мозге. Начало и конец рефлекторной дуги связаны
с мышцей. Рефлексы наиболее выражены в мышцах—разгибателях. Для того чтобы
организм мог противостоять силе земного притяжения, эти мышцы должны находиться
в состоянии тонического напряжения. Биологическое значение этих рефлексов
состоит в том, что они участвуют в сохранении статики и положения тела,
регулируя степень сокращения мышцы в соответствии с действующими на нее
раздражениями. Особое значение этот вид рефлексов имеет у копытных животных,
хотя и у человека они хорошо развиты.
|
|
|
Рис. 3.16
Дуга рефлекса растяжения и реципрокного торможения мышц—антагонистов С —
мотонейроны сгибателей коленного сустава; Р — мотонейроны разгибателей
коленного сустава. 1 — волокна Iа, 2 — двигательные
аксоны, 3 — концевые пластинки, 4 — сгибатель, 5 — разгибатель. |
Рефлексы мышц—антагонистов (рис. 3.17) лежат в основе
локомоторных актов и характеризуются тем, что при возбуждении мотонейронов сгибателей
одновременно происходит торможение мотонейронов мыщц—разгибателей. При этом в
конечности другой стороны наблюдаются обратные явления. В целом это создает
правильное чередование противоположных по функциональному значению мышечных
сокращений. Механизм, обусловливающий такое чередование активности различных
двигательных ядер, например при ходьбе, локализуется в интернейронном аппарате
спинного мозга. Вместе с тем для его активации необходимо поступление
тонического нисходящего сигнала из двигательных центров головного мозга.
Висцеромоторные рефлексы возникают при возбуждении
афферентных волокон внутренних органов и характеризуются появлением
двигательных реакций мышц грудной и брюшной стенки, мышц—разгибателей спины.
Возникновение этих рефлексов связано с существованием конвергенции висцеральных
и соматических афферентных волокон к одним и тем же интернейронам спинного
мозга.
Вегетативные рефлексы заключаются, во—первых, в появлении
полисинаптических разрядов в преганглионарных симпатических волокнах в ответ на
возбуждение
|
|
|
Рис. 3.17 Нервные пути сгибательного и разгибательного рефлекса С —
ипсилатеральные сгибательные мотонейроны; Р — ипсилатеральные разгибательные
мотонейроны. 1 — белое
вещество, 2 — разгибатель, 3
— сгибатель. |
|
|
|
Рис. 3.18 Соединение кожных и висцеральных афферентное в спинном мозге I — кожа, 2 — механорецепторы, 3 — нисходящие пути, 4 — тормозный нейрон, 5 —
дорсальный столб, 6 — поясничный
отдел, 7 — грудной отдел, 8 — шейный
отдел, 9 — вентролатеральный
канатик, 10 — дорсальный
корешок, II —
вентральный корешок, 12 —
проприоспинальный путь, 13 —
двигательные аксоны, 14 —
симпатические аксоны, 15 —
вентральный рог, 16 — внутренности,
17 — висперопепторы, 18 — мышца, 19 — проприопепторы, 20
— дорсальный рог, 21 —
терморецепторы, 22 —
ноцицепторы. |
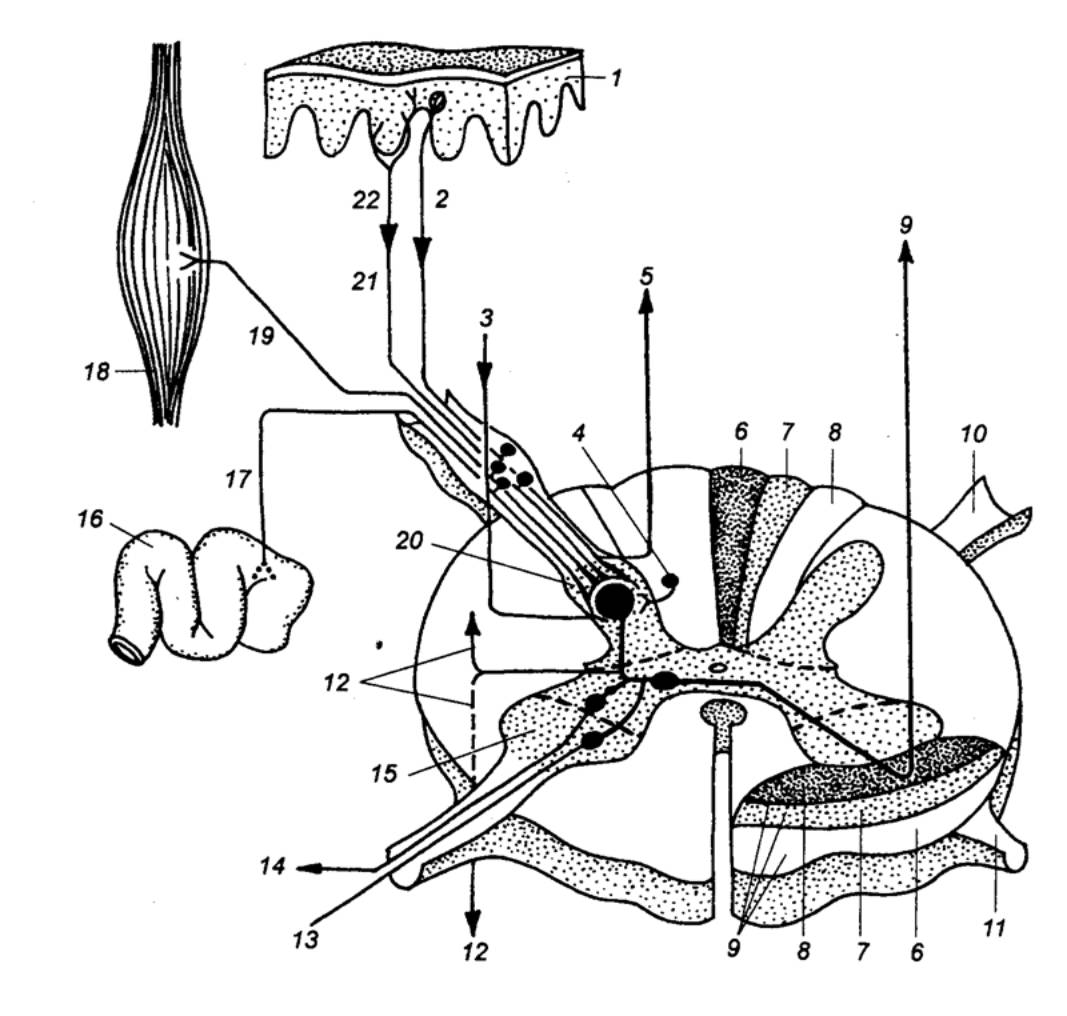
симпатических и соматических
чувствительных клеток (рис. 3.18), во—вторых, в возникновении рефлекторных
реакций парасимпатических нейронов в ответ на раздражение чувствительных путей (более
подробно эти реакции рассмотрены в разд. 5.3).
Наряду с выполнением собственных рефлекторных реакций
нейронные структуры спинного мозга служат аппаратом для реализации большого
числа сложных процессов, осуществляемых различными отделами головного мозга.
Это управление может быть прямым, когда нисходящие пути непосредственно связаны
с мотонейронами спинного мозга, и непрямым через интернейроны образующие
короткие межсегментарные связи. Свойства последних и особенности связей с
нисходящими волокнами и моторными нейронами создают возможность к интеграции
поступающих сигналов, их переработке и пространственному перераспределению.
3.3.3. Проводниковые функции спинного мозга
Помимо рефлекторной деятельности еще одной важной функцией
спинного мозга является проведение импульсов. Оно осуществляется белым
веществом, состоящим из нервных волокон.
В результате эволюционного развития простая рефлекторная
дуга, лежащая в основе функции нервной системы, усложняется, и в каждой ее
части вместо одного нейрона возникают цепи нервных клеток, аксоны которых
образуют проводящие пути. Под проводящими путями принято понимать группы
Таблица 3.2
Основные проводящие пути спинного мозга
|
Проводящие
пути |
Столбы
спинного мозга |
Физиологическое
значение |
|
А.
Восходящие (чувствительные) пути |
||
|
1.
Тонкий пучок (пучок Голля) |
Дорсальные |
Тактильная
чувствительность, чувства положения тела, пассивных движений тела, |
|
2.Клиновидный
(пучок Бурдаха) |
« |
Тоже |
|
3.
Дорсолатеральный |
Боковые |
Пути
болевой и температурной чувствительности |
|
4.
Дорсальный спинно—мозжечковый путь (пучок Флексига) |
« |
Импульсы
от проприоцепторов мышц, сухожилий, связок; чувство давления и прикосновения
из кожи |
|
5.
Вентральный спинно—мозжечковый путь (пучок Говерса) |
« |
Тоже |
|
таламический
путь |
« |
Болевая
и температурная чувствительность |
|
7.
Спинно—тектальный путь таламический путь |
« |
Сенсорные
пути зрительно—двигательных рефлексов (?) и болевой чувствительности (?) |
|
Вентральные |
Тактильная
чувствительность |
|
|
|
||
|
1.
Латеральный корково—спинномозговой (пирамидный) путь |
Боковые |
Импульсы
к скелетным мышцам. Произвольные движения |
|
2.
Красноядерно—спинномозговой (Монакова) путь |
« |
Импульсы,
поддерживающие тонус скелетных мышц |
|
спинномозговой
путь |
« |
позы и
равновесия тела |
|
4.
Оливоспинномозговой (Гельвёга) путь ' |
« |
Функция неизвестна.
Возможно, он участвует в осуществлении таламоспинальных рефлексов |
|
5.
Ретикулярно—спинномозговой путь |
Вентральные |
Импульсы,
поддерживающие тонус скелетных мышц, регулирующие состояние спинальных
вегетативных центров и чувствительность мышечных веретен проприорецепторов
скелетных мышц |
|
6.
Вентральный преддверно—спинномозговой путь |
« |
Импульсы,
обеспечивающие поддержание позы и равновесия тела |
|
7.
Тектоспинальный (покрышечно—спинномозговой) путь |
« |
Импульсы,
обеспечивающие осуществление зрительных и слуховых двигательных рефлексов |
|
8.
Вентральный корково—спинномозговой (пирамидный) путь |
Вентральные |
Импульсы
к скелетным мышцам, произвольные движения |
нервных волокон, характеризующиеся общностью
строения и функций. Они связывают различные отделы спинного мозга или спинной и
головной мозг (табл. 3.2). Все нервные волокна одного пути начинаются от
однородных нейронов и заканчиваются на нейронах, выполняющих одинаковую
функцию.
В соответствии с функциональными особенностями различают
ассоциативные, комиссуральные и проекционные (афферентные и эфферентные)
нервные волокна. Ассоциативные волокна, или пучки осуществляют
односторонние связи между отдельными частями спинного мозга. Связывая разные
сегменты, они образуют собственные пучки, являющиеся частью сегментарного
аппарата спинного мозга. Комиссуральные волокна соединяют функционально
однородные противоположные участки разных отделов спинного мозга. Проекционные
волокна связывают спинной мозг с вышележащими отделами. Эти волокна
образуют основные проводящие пути, которые представлены восходящими
(центростремительными, афферентными, чувствительными) и нисходящими
(центробежными, эфферентными, двигательными) путями.
Восходящие проводящие пути. Несут импульсы от рецепторов,
воспринимающих информацию из внешнего мира и внутренней среды организма. В
зависимости от вида чувствительности, которую они проводят, их делят на пути экстеро—,
проприо— и интероцептивной чувствительности. Нисходящие пути передают
импульсы от структур головного мозга к двигательным ядрам, осуществляющим
ответные реакции на внешние и внутренние раздражения.
Основными восходящими путями спинного мозга являются тонкий
пучок, клиновидный пучок, латеральный и вентральный спинно—таламические пути,
дорсальный и вентральный спинно—мозжечковые пути (рис. 3.19).
Тонкий пучок (Голля) и клиновидный пучок (Бурдаха) составляют
задние канатики спинного мозга. Эти пучки волокон являются отростками
чувствительных клеток спинальных ганглиев, проводящих возбуждение от
проприорецепторов мышц, сухожилий, частично тактильных рецепторов кожи,
висцерорецепторов. Волокна тонкого и клиновидного пучков — миелинизированные,
они проводят возбуждение со скоростью 60—100 м/с. Короткие аксоны обоих пучков
устанавливают синаптические связи с мотонейронами и интернейронами своего
сегмента, длинные же направляются в продолговатый мозг. По пути они отдают
большое число ветвей к нейронам вышележащих сегментов спинного мозга, образуя,
таким образом, межсегментарные связи.
По волокнам тонкого пучка проводится возбуждение от
каудальной части тела и тазовых конечностей, по волокнам клиновидного пучка —
от краниальной части тела и грудных конечностей. В спинном мозгу оба этих пути
идут, не прерываясь и не перекрещиваясь, и оканчиваются в продолговатом мозгу у
одноименных ядер, где образуют синаптическое переключение на второй нейрон.
Отростки второго нейрона направляются к специфическим ядрам таламуса
противоположной стороны, образуя тем самым своеобразный перекрест. Здесь
они переключаются уже на третий нейрон, аксоны которого достигают нейронов IV
слоя коры больших полушарий.
Считают, что по этой системе проводится информация тонко
дифференцированной чувствительности, позволяющая определить локализацию, контур
периферического раздражения, а также его изменения во времени.
По латеральному спинно—таламическому пути проводится
болевая и температурная чувствительность, по вентральному
спинно—таламическому — тактильная. Существуют сведения, что по этим путям возможна
также передача возбуждения от проприо— и висцероцепторов. Скорость проведения
возбуждения в волокнах составляет 1—30 м/с. Спинно—таламические пути
прерываются
|
|
|
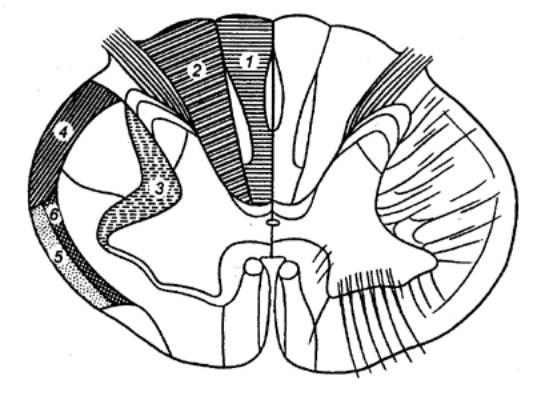
Рис. 3.19 Локализация основных восходящих путей в белом
веществе спинного мозга 1— тонкий пучок (Голля), 2— клиновидный пучок (Бурдаха), 3
— дорсолатеральный, 4 — дорсальный спинномозжечковый, 5 — вентральный
спинномозжечковый (Говерса), 6 — дорсальный спинио—таламический. |
|
|
|
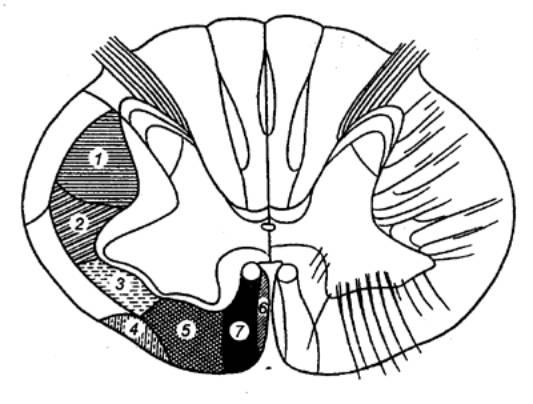
Рис. 3.20 Локализация основных нисходящих
путей спинного мозга 1 — латеральный корково—спинномозговой (пирамидный), 2 — красноядерно—спинномозговой (Монакова), 3 — дорсальный
преддверно—спинномозговой, 4 — оливоспинномозговой (Гельвега), 5 —
вентральный преддверно—спинномозговой, 6 — покрышечно—спинномозговой, 7 —
вентральный корково—спинномозговой (пирамидный). |
и перекрещиваются либо на уровне
сегмента, в который они только что вступили, либо вначале проходят несколько сегментов
по своей стороне, а затем переходят на противоположную. Отсюда идут волокна,
оканчивающиеся в таламусе. Там они образуют синапсы на нервных клетках, аксоны
которых направляются в кору больших полушарий.
Полагают, что по системе волокон этих путей в основном
передается информация о качественной природе раздражителей.
Дорсальный спинно—мозжечковый путь, или пучок Флексига —
филогенетически это наиболее древний чувствительный путь спинного мозга. Местом
расположения нервных клеток, аксоны которых образуют волокна этого пути,
является основание дорсального рога спинного мозга. Не перекрещиваясь, путь
достигает мозжечка, где каждое волокно занимает определенную область. Скорость
проведения по волокнам спинно—мозжечкового пути около 110 м/с. По ним проводится
информация от рецепторов мышц и связок конечностей. Наибольшего развития этот
путь достигает у копытных животных.
Вентральный спинно—мозжечковый путь, или пучок Говерса, также образуется
аксонами интернейронов противоположной стороны спинного мозга. Через
продолговатый мозг и ножки мозжечка волокна направляются к коре мозжечка, где
занимают обширные площади. Импульсы со скоростью проведения до 120 м/с идут от
сухожильных, кожных и висцерорецепторов. Они участвуют в поддержании тонуса
мышц для выполнения движений и сохранения позы.
Нисходящие проводящие пути. Эти пути (рис. 3.20) связывают
высшие отделы ЦНС с эффекторными нейронами спинного мозга. Основными из них
являются пирамидный, красноядерно—спинномозговой и ретикулярно—спинномозговой
пути.
Пирамидный путь образован аксонами клеток двигательной зоны коры больших
полушарий. Направляясь к продолговатому мозгу, эти аксоны отдают большое число
коллатералей структурам промежуточного, среднего, продолговатого мозга и
ретикулярной формации. В нижней части продолговатого мозга большая часть
волокон пирамидного пути переходит на противоположную сторону (перекрест
пирамид), образуя латеральный пирамидный путь. В спинном
мозгу он располагается в боковом
канатике. Другая часть волокон идет, не перекрещиваясь, до спинного мозга и
только на уровне сегмента, в котором оканчивается, переходит на противоположную
сторону. Это прямой вентральный пирамидный путь. Оба заканчиваются на
мотонейронах передних рогов серого вещества спинного мозга. Состав волокон
этого пути неоднороден, в нем представлены миелинизированные и
немиелинизированные волокна разного диаметра со скоростями проведения
возбуждения от 1 до 100 м/с.
Основной функцией пирамидных путей является передача импульсов для
выполнения произвольных движений. Надежность в осуществлении этой функции
повышается благодаря дублированию связи головного мозга со спинным посредством
двух путей — перекрещенного и прямого. В эволюционном ряду пирамидный тракт
развивался параллельно с развитием коры больших полушарий и достиг наибольшего
совершенства у человека.
Красноядерно—спинномозговой путь (Монакова) образован аксонами
клеток красного ядра среднего мозга. Выйдя из ядра, волокна полностью переходят
на противоположную сторону. Часть из них направляется в мозжечок и ретикулярную
формацию, другие — в спинной мозг. В спинном мозгу волокна располагаются в
боковых столбах перед перекрещенным пирамидным путем и оканчиваются на
интернейронах соответствующих сегментов. Красноядерно—спинномозговой путь несет
импульсы от мозжечка, ядра вестибулярного нерва, полосатого тела.
Основное назначение
красноядерно—спинномозгового пути — управление тонусом мышц и непроизвольной
координацией движений. В процессе эволюции этот путь возник рано. Большое
значение он имеет у животных, слабее развит у человека.
Преддверно—спинномозговой путь образован волокнами, которые
являются отростками клеток латерального пред дверного ядра (ядра Дейтерса),
лежащего в продолговатом мозгу. Этот тракт имеет наиболее древнее эволюционное
происхождение. По нему передаются импульсы от вестибулярного аппарата и
мозжечка к мотонейронам вентральных рогов спинного мозга, регулирующие тонус
мускулатуры, согласованность движений, равновесие. При нарушении целостности
этого пути наблюдаются расстройства координации движений и ориентации в
пространстве.
В спинном мозгу помимо основных
длинных имеются и короткие нисходящие пути, соединяющие между собой его
отдельные сегменты.