13.6.7. Роль почек в осморегуляции и волюморегуляции
Осморегуляция. Почки являются основным эффекторным
органом системы осморегуляции. Они обеспечивают выделение избытка воды в виде
гипотонической мочи при поступлении воды в организм или экономят воду и экскретируют
мочу, гипертоническую по отношению к плазме крови, при обезвоживании. При
избыточном содержании воды в организме концентрация растворенных осмотически
активных веществ в крови снижается и ее осмотическое давление падает. Это
уменьшает активность центральных осморецепторов, расположенных в области
супраоптического ядра гипоталамуса, а также периферических осморецепторов,
имеющихся в печени, почке, селезенке и ряде других органов. Уменьшение
активности осморецепторов снижает секрецию антидиуретического гормона (АДГ)
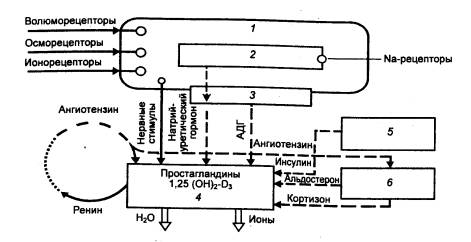
нейрогипофизом и приводит к возрастанию выделения воды почкой (рис. 13.14).
|
|
|
Рис.
13.14 Участие почки в регуляции
водно—солевого обмена 1— ЦНС, 2 — гипоталамус, 3 — нейрогипофиз, 4 — почка, 5 — поджелудочная
железа, 6 — надпочечник; |
При обезвоживании организма, при
введении в сосудистое русло гипертонического раствора хлористого натрия
увеличивается концентрация осмотически активных веществ в плазме крови,
возбуждаются осморецепторы, стимулируются нейроны супраоптического ядра,
усиливается секреция АДГ, возрастает реабсорбция воды в канальцах, уменьшается
мочеотделение и выделяется осмотически концентрированная моча. Установлено, что
у человека в норме при осмоляльности крови около 280 мосм/кг H2O полностью прекращается секреция АДГ
и почка выделяет наибольшие количества воды. При обезвоживании когда
осмоляльность крови возрастает до 295 мосм/кг Н2О, секреция АДГ
достигает максимума и обнаруживается пик осмотического концентрирования мочи в
почке. В эксперименте показано, что секреция АДГ возрастает при раздражении не
только осморецепторов, но и специфических натриорецепторов. После
введения в область III желудочка мозга гипертонического раствора NaСl наступает антидиурез, а после
введения в ту же область гипертонических растворов сахара мочеотделение не
уменьшается.
Осморецепторы реагируют на сдвиги
концентрации осмотически активных веществ в плазме крови. При увеличении
осмоляльности плазмы крови на 1% (около 3 мосм/л) концентрация
аргинин—вазопрессина, являющегося антидиуретическим гормоном у человека и
многих других млекопитающих, возрастает на 1 пг/мл плазмы. Повышение
концентрации осмотически активных веществ в плазме на 1 мосм/кг воды вызывает
благодаря выделению АДГ увеличение осмотической концентрации мочи почти на 100
мосм/кг H2O, а переход от состояния водного диуреза до максимального осмотического
концентрирования мочи требует всего 10—кратного возрастания АДГ в крови — с 0,5
до 5 пг/мл.
Волюморецепция. Помимо информации от осморецепторов
и натриорецепторов уровень секреции АДГ зависит и от активности волюморецепторов,
реагирующих на изменение объема внутрисосудистой и внеклеточной жидкости.
Ведущее значение в регуляции секреции АДГ имеют те волюморецепторы, которые
реагируют на изменение напряжения сосудистой стенки. В первую очередь это
рецепторы левого предсердия, импульсы от которых передаются в ЦНС по
афферентным волокнам блуждающего нерва. При увеличении кровенаполнения левого
предсердия активируются волюморецепторы и угнетается секреция АДГ, тем самым
усиливается мочеотделение. В предсердии вырабатывается гормон, увеличивающий
выделение ионов Na+ почкой и получивший название атриальный
натрийуретический пептид.
Рассмотрим особенности реакции почки
при возбуждении волюморецепторов и осморецепторов. Активация только
волюморецепторов возникает в ответ на увеличение объема внутрисосудистой
жидкости при неизмененной концентрации в ней осмотически активных веществ. Это
приводит к повышению экскреции солей натрия и воды. Одновременная активация
волюморецепторов и осморецепторов может наступить при потреблении больших
количеств воды, в этом случае в основном усиливается выведение воды. Ведущую
роль в изолированном осморегулирующем рефлексе играет уменьшение секреции АДГ,
в результате снижается реабсорбция воды и усиливается ее экскреция.
Увеличение объема внеклеточной
жидкости, возрастание объема крови служат стимулом для волюморегулирующего
рефлекса. В эксперименте адекватным раздражителем системы регуляции объема
внеклеточной жидкости может служить введение в кровеносный сосуд больших
количеств изотоничного плазме крови физиологического раствора. Возрастание
притока крови к сердцу, растяжение стенки предсердия стимулируют
волюморецепторы и секрецию из клеток предсердия атриального натрийуретического
пептида. Под влиянием этого гормона усиливается выделение ионов натрия и воды
почкой.
Стимуляция волюморецепторов
рефлекторно изменяет характер импульсации иннервирующих почку симпатических
нервов и снижает реабсорбцию натрия и воды в канальцах. Возрастание объема
крови способствует растяжению афферентной артериолы клубочка. Это
сопровождается изменением активности юкстагломерулярного аппарата, уменьшением
секреции фермента ренина, благодаря чему снижается образование в крови из
ангиотензиногена ангиотензина I. После отщепления от него двух
аминокислот образуется ангиотензин II, который вызывает ряд эффектов, в
том числе стимулирует секрецию альдостерона. Когда снижается количество
ангиотензина, падает и поступление в кровь альдостерона, что приводит к
уменьшению реабсорбции натрия в канальцах. Активация волюморегулирующего
рефлекса снижает секрецию вазопрессина, тем самым уменьшается
реабсорбция воды и повышается ее выделение почкой.
Клеточное действие вазопрессина. Воздействие вазопрессина начинается
с активации рецепторов, расположенных на базальной и латеральных мембранах
клеток конечных частей дистального сегмента нефрона и собирательных трубок.
Существует два типа рецепторов вазопрессина: V1 и V2. Первый тип рецепторов после
взаимодействия с вазопрессином активирует образование таких вторичных
мессенджеров, как инозитолтрифосфат и диацилглицерол. Стимуляция
вазопрессином V2—рецепторов
завершается активацией фермента аденилатциклазы, в результате из АТФ
образуется вторичный мессенджер — цАМФ. Это физиологически активное
вещество внутри клетки вызывает ряд превращений, приводящих к увеличению
проницаемости для воды апикальной (люминальной) плазматической мембраны,
обращенной в просвет канальца.
В обычных условиях, когда в крови
нет вазопрессина, люминальная мембрана непроницаема для воды, клетка не
пропускает через себя воду и реабсорбция воды в собирательных трубках
практически не происходит. После секреции вазопрессина в кровь и стимуляции им
V2—рецепторов у внутренней поверхности базальной плазматической
мембраны образуется цАМФ. Этот вторичный мессенджер должен пересечь клетку и у
противоположной плазматической мембраны активировать ряд процессов,
завершающихся увеличением проницаемости для воды.
В цитоплазме клетки содержатся агрефоры.
— в них находятся так называемые «водные каналы» (аквапорины), которые
перемещаются к люминальной мембране и встраиваются в нее. Для осуществления
этого процесса необходимо участие микрофиламентов и микротрубочек. В конечном
счете цАМФ способствует встраиванию в люминальную мембрану агрегатов
внутримембранных частиц;
чем больше образуется цАМФ, тем
большая поверхность мембраны будет занята агрегатами частиц и выше становится
проницаемость для воды. Восстановление исходной проницаемости зависит от
скорости ферментативного разрушения цАМФ, эту функцию выполняет имеющаяся
внутри клетки фосфодиэстераза цАМФ.
Таким образом, активация V2—рецепторов вазопрессином сопровождается
образованием цАМФ и увеличением проницаемости для воды.
Выше речь шла о V1—рецепторах. Их физиологическое
значение состоит в модуляции, регуляции силы ответа клетки, они обеспечивают
осуществление своеобразной системы обратной связи. При регуляции водного обмена
в организме ведущую роль играет лишь один гормон — вазопрессин.
В регуляции ионного обмена обычно
природа использует два гормона, оказывающие противоположное физиологическое
действие, например кальцитонин, снижающий концентрацию кальция в крови,
и паратгормон, увеличивающий ее.
В случае вазопрессина можно описать уникальный
способ физиологической регуляции, когда один гормон с помощью рецепторов
двух разных типов стимулирует реакцию и модифицирует ее. Активация V1—рецепторов и образование описанных
выше вторичных мессенджеров способствуют снижению силы действия цАМФ.
Экспериментально можно активировать только аденилатциклазу и устранить
фосфоинозитидный ответ, связанный со стимуляцией V1—рецепторов. В
таком случае сила ответа на вазопрессин станет выше.
Эти данные позволяют представить
целостную картину регуляции водного обмена: от возбуждения осморецепторов и
секреции вазопрессина до конечных этапов внутриклеточных изменений,
сопровождающихся возрастанием проницаемости мембраны для воды. Поток воды при
увеличении ее реабсорбции растет в десятки раз.
Выше шла речь о процессах,
происходящих в базальной мембране, цитоплазме и апикальной мембране. Такие
большие физиологические изменения, сопровождающиеся возрастанием потока воды,
затрагивают и внешнюю поверхность клетки. После добавления вазопрессина наблюдается
экзоцитоз — выделение веществ со стороны апикальной мембраны на ее
внешнюю поверхность. Среди этих веществ находятся компоненты гликокаликса, а
также, вероятно, и ряд гидролитических ферментов.
А. Г. Гинецинский высказал предположение,
что на одном из этапов действия АДГ в процесс вовлекается гиалуронатгидролаза,
деполимеризующая гликозаминогликаны в межклеточном веществе почки. В пользу
такого предположения свидетельствуют данные, что введение в кровь антисыворотки
к почечной гиалуронатгидролазе уменьшает способность АДГ увеличивать
проницаемость для воды и ее реабсорбцию.