10.1.2. Дыхание позвоночных животных
Все позвоночные обладают
специализированными органами дыхания (за исключением некоторых амфибий,
вторично утративших их). Эти органы, как правило, развиваются из выростов
кишечной трубки (выше было указано на филогенетическую связь функций дыхания и
питания).
Общий принцип функционирования респираторных систем
заключается в создании динамического контакта крови с газовой средой, в
сочетании кровотока в сосудах органов дыхания (их перфузии) с прокачиванием
через эти органы воды или воздуха (их вентиляции). Исключение составляет кожное
дыхание амфибий, не требующее особого вентилирующего аппарата.
Водное дыхание. Водное дыхание с помощью жабр присуще большинству рыб. Эти
животные набирают воду в ротовую полость, после чего вода выталкивается через
жаберные щели. Для извлечения кислорода используется принцип противотока
(рис. 10.4). Кровь в жаберных капиллярах течет навстречу воде, омывающей
жаберные листки, чему способствует перемещение рыбы в пространстве. Высокая
эффективность жаберного дыхания призвана компенсировать большие энерготраты на
работу мускулатуры, которая осуществляет вентиляцию жабр водой. Этой
мускулатурой управляет обособленный дыхательный центр
|
|
|
Рис. 10.4. Жаберные
дуги (четыре) у рыбы. (А), фрагмент двух жаберных дуг (Б), участок жаберной
нити с тремя жаберными лепестками на каждой стороне (В), а также участок жабр
костистой рыбы., в которых кровь и вода текут в противоположных направлениях
(Г) 1 — жаберная крышка, 2
— констрикторная мышца, 3 — жаберные нити, 4 — жаберная полость, 5 — кровеносные сосуды, 6 — ротовая полость, 7 — жаберный скелет, 8 — пластины, 9 — поперечный срез через жаберную
нить, 10 — перегородка, 11 — перегородочный канал, 12 — край пластины. |

в продолговатом мозгу, приближающийся по своей
организации к дыхательному центру млекопитающих. Открытие ритмической
активности структур изолированного ствола мозга золотой рыбки (Carassius auratus auratus) явилось важной вехой в познании автоматизма дыхательного
центра.
В связи с большой растворимостью СО2
в воде отдача его происходит легко, поэтому работа респираторного аппарата рыбы
служит в основном обеспечению организма кислородом. Сигналы о содержании О2
в крови, поступающие от хеморецепторов жаберных сосудов, не только регулируют
активность дыхательного центра рыбы, но и побуждают ее выбирать зоны водоема с
физиологически оптимальной оксигенацией. Подобную поведенческую реакцию
активного выбора предпочитаемой дыхательной среды называют газопреферендумом.
Некоторые рыбы (угри) поглощают растворенный в воде кислород преимущественно не
через жабры, а используя его диффузию через кожу.
Переход от водного дыхания к
воздушному. Многие
рыбы, спорадически поднимаясь на поверхность водоемов или даже выходя на сушу и
заглатывая воздух, дополняют водное дыхание воздушным, для чего используют
богато васкуляризированные стенки ротовой полости, глотки, желудка, кишки либо
особый дериват кишечной трубки — плавательный пузырь.
Двоякодышащие рыбы. дышат воздухом уже с помощью нового
специализированного органа — легкого, которое представляет собой
складчатый мешочек (одиночный или парный), происходящий из вентрального
выпячивания глотки. Немногочисленные виды двулегочникообразных рыб (Lepidosireniformes) обитают в тропических водах, плохо аэрируемых и к
тому же подкисленных. Они не могут обходиться без воздушного дыхания.
|
|
|
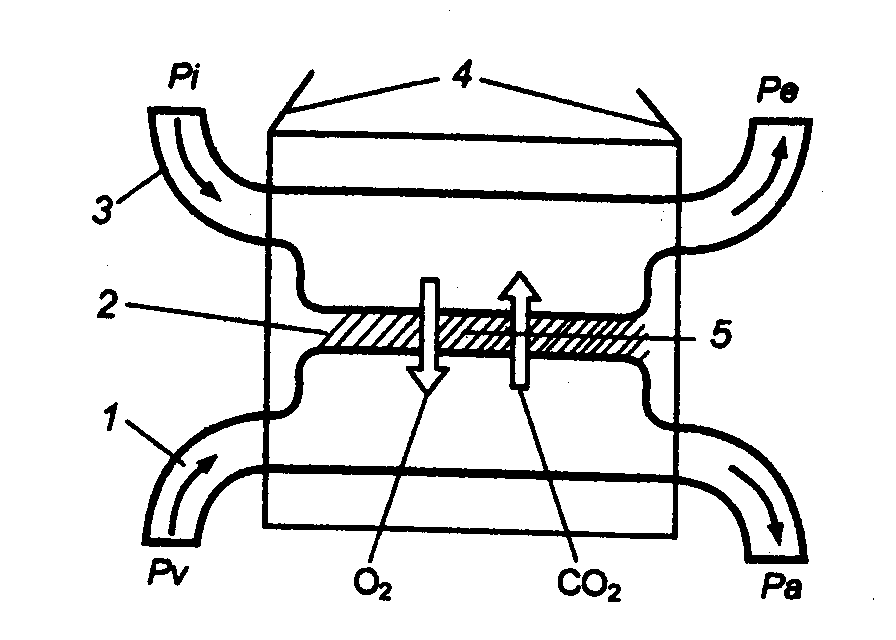
Рис.
10.5 Принцип функционирования
респираторных систем позвоночных 1 — кровь, 2 — барьер, 3 — среда, 4
— органы дыхания, 5 — диффузия; Pi, Ре —
парциальные давления О2 во вдыхаемом и выдыхаемом воздухе (воде); Pa, Pv — то же в
артериальной и венозной крови. |
Подобное изменение условий,
очевидно, и способствовало массовому переходу водных позвоночных (это были
кистеперые рыбы — предки амфибий) к воздушному дыханию в девонский период
(около 400 млн. лет тому назад). Следует учесть, что в ту эпоху парциальное
давление 02 в атмосфере было близко к современному. И если в 1 л воды, насыщенной
воздухом при температуре О °С, содержится всего 0,014 г 02 (при 30 °С это
количество уменьшается вдвое), то в 1 л воздуха его содержание составляет 0,28
г, т. е. в 20 раз больше. Кроме того, плотность воздуха более чем в 700 раз
ниже, чем воды, поэтому его вентиляция через органы дыхания требует
соответственно меньших энерготрат. На дыхание воздухом перешло большинство
амфибий (во всяком случае их взрослые формы), хотя легкие земноводных развиты
еще слабо: газообменная поверхность невелика, потоки венозной и артериальной
крови в системе циркуляции разведены не полностью. Некоторая доля поглощаемого
О2 и большая часть выделяемого СО2 диффундирует не через
легкие, а через увлажненную кожу, васкуляризация которой меняется в
соответствии с интенсивностью кожного газообмена (рис. 10.5).
Дыхательные движения амфибий
(как и рептилий) характерны — этим они отличаются и от рыб, и от млекопитающих
— длительными остановками, апноэ, во время которых вентилируется только щечно—глоточная
полость при закрытой голосовой щели, отделяющей легкие от атмосферы. Лишь время
от времени возникают координированные сокращения мышц рта и гортани,
нагнетающие воздух в легкие и отсасывающие его обратно. Эти движения
управляются нервными центрами ствола мозга в соответствии с импульсацией
рецепторов растяжения легких, а также артериальных хеморецепторов, возбуждаемых
недостатком кислорода в крови. Кроме того, у многих амфибий наблюдается
поведенческая регуляция дыхания, газопреферендум: в зависимости от условий они
избирают либо водную, либо воздушную среду.
У наземных рептилий в связи с
развитием ороговевающего покрова водное и кожное дыхание становится невозможным
(за исключением ящериц). Данный этап эволюции дыхания знаменует окончательный
переход от газообмена во влажной среде к «сухой» вентиляции, механизм которой
уже почти такой же, как у млекопитающих. Газообменная поверхность легких
значительно увеличивается за счет образования мелких ячеек, через густую сеть
их сосудов протекает кровь уже обособившегося малого круга. Вместе с тем
рептилии еще сохраняют прерывистый характер вентиляции легких, которые
заполняются воздухом, однако путем не нагнетания его, а засасывания.
Метаболизм эктотермных
(пойкилотермных) позвоночных еще относительно низок, с чем связана
сравнительно малая «пропускная способность» их дыхательного аппарата. Как
полагают некоторые исследователи, это и явилось причиной неспособности ныне
вымерших крупных рептилий адаптироваться к происшедшим в конце мезозоя резким
изменениям в среде обитания.
Значительно интенсивнее и устойчивее
обменные процессы у эндотермов (гомойотермных животных) — млекопитающих
и птиц. Происходит дальнейшее увеличение газообменной поверхности и
кровоснабжения легких. В отличие от амфибий и рептилий вновь формируется ритмичный,
без пауз, тип вентиляции легких. Существенно изменяется механизм регуляции
дыхания из—за того, что весь CO2 у этих животных удаляется
практически только легкими, а продукция его становится в связи с усилением окислительных
процессов весьма интенсивной. Возникает затруднение в борьбе с накоплением
угольной кислоты в тканях и крови. Поэтому у животных, дышащих исключительно
легкими, хотя и увеличивается Роз артериальной крови, одновременно значительно
возрастает Рсо2 (рис. 10.6), усиливается и мощность буферных систем.
|
|
|
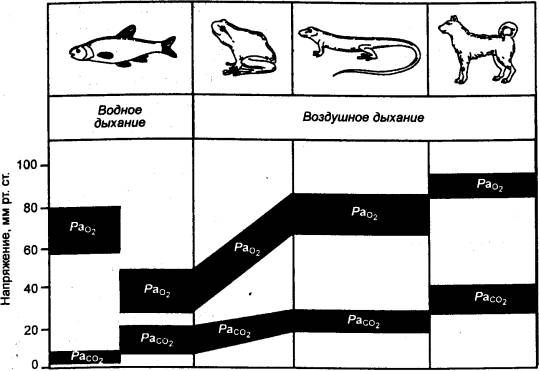
Рис. 10.6 Напряжение дыхательных газов в артериальной крови (РаО2 Pа co2) у
различных классов позвоночных Жаберное
дыхание рыб обеспечивает их кислородом; Р co2 крови у них низкое, поскольку co2легко диффундирует через покровы
тела в окружающую воду. Дыхательное поведение и жаберное дыхание рыб
регулируются по кислороду. У тех рыб, которые в определенных условиях
используют воздушное дыхание (а также у рептилий и млекопитающих), не только
поступление О2, но и отдача co2 в воздушную среду происходят
только через легкие. Поэтому Р co2 крови у воздушнодышащих животных
гораздо выше, чем у воднодышащих, и этот параметр становится главным
регулятором дыхания. Вместе с тем Рo2 крови у водно— и воздушнодышащих
различаются незначительно. |
В силу данных обстоятельств роль
главного регулятора дыхания переходит от О2 к СО2 и
сопряженным с его транспортом ионам Н+, а ведущее значение в
стимуляции ритмической активности дыхательного центра приобретают вместо
артериальных хеморецепторов центральные (медуллярные) хемочувствительные
зоны. Все эти особенности будут освещены в соответствующих разделах главы.
Вместе с тем дыхание птиц настолько отличается от дыхания млекопитающих, что на
нем следует остановиться особо.
Дыхание птиц. Легкие птиц, имеющие губчатое
строение, отличаются, во—первых, тем, что газообмен между воздухом и кровью
происходит в так называемых воздушных капиллярах, «цилиндрических альвеолах»
(по аналогии с мешковидными альвеолами млекопитающих) — и, во—вторых, проточной
системой бронхов. Эта система устроена таким образом, что легкие с помощью
нескольких воздушных мешков вентилируются свежим воздухом и при вдохе, и
при выдохе (см. рис. 10.7). Такой тип дыхания придает легочному газообмену
птицы высокую эффективность, необходимую для покрытия интенсивного расходования
энергии, особенно в полете. Дыхательные движения грудной стенки создаются в
основном сокращениями грудных и брюшных мышц. В полете вентиляции легких
(которая помимо газообмена участвует в отдаче избыточного тепла) содействуют
движения крыльев.
Дыхательный центр птицы расположен в
продолговатом мозгу. Его активность, как и у млекопитающих, контролируется
импульсацией от медуллярных хеморецепторов, чувствительных к СО2 и
ионам Н+, а также артериальных хеморецепторов, реагирующих, кроме
того, и на недостаток О2, что имеет значение для регуляции дыхания
во время полета на больших высотах. В воздухоносных путях птицы находятся механорецепторы,
стимуляция которых участвует в регуляции частоты и глубины дыхания. Там же
имеются особые хеморецепторы, импульсация из которых усиливается при снижении
внутрилегочной концентрации CO2. Стимуляция этих рецепторов
угнетает активность дыхательного центра и снижает тем самым вентиляцию легких,
предотвращая чрезмерное вымывание СО2 из организма, которое могло бы
происходить в условиях полета.
|
|
|
Рис. 10.7 Расположение воздушных мешков и легких у птиц
(А), а также схема строения легкого птицы, (Б), бронхиального дерева и
соединенных с ним воздушных мешков (В) |
