9.6.2.
Нейрогуморальная регуляция системного кровообращения
Координационные акты, которые приспосабливают
сердечно—сосудистую систему к оптимальному обеспечению кровоснабжением органов
и тканей в соответствии с их функциональным состоянием, осуществляются за счет
деятельности нервной системы и гуморальных факторов. Эта регуляция
обеспечивается сложным механизмом, включающим чувствительное, центральное и
эфферентное звенья. Эфферентное звено представлено нервным и эндокринным
компонентами.
Чувствительное звено. Чувствительная иннервация сосудов представлена
главным образом свободными древовидно разветвляющимися нервными окончаниями,
которые располагаются между коллагеновыми волокнами адвентиции. Рецепторы
сосудов — ангиоцепторы — по своей функции подразделяются на барорецепторы
(прессорецепторы), реагирующие на изменение артериального давления, и хеморецепторы,
чувствительные к изменению химического состава крови. Раздражителем
барорецепторов является не давление как таковое, а скорость и степень
растяжения стенки сосуда пульсовыми или нарастающими колебаниями кровяного
давления.
Ангиоцепторы расположены по всей сосудистой системе и
составляют единое рецептивное поле (В. Н. Черниговский). Их наибольшие
скопления находятся в главных рефлексогенных зонах: аортальной,
синокаротидной, в сосудах легочного круга кровообращения. В ответ на каждое
систолическое повышение артериального давления барорецепторы зон генерируют
залп импульсов, которые затухают при диастолическом снижении давления.
Минимальный порог возбуждения барорецепторов — 40 мм рт. ст., максимальный —
200 мм рт. ст. Повышение давления выше этого уровня не ведет к дополнительному
учащению импульсации.
Аортальная рефлексогенная зона. Существование этой зоны было открыто И. Ф. Ционом и
К. Людвигом в 1866 г. При повышении артериального давления происходит
растяжение стенки аорты, что влечет за собой раздражение барорецепторов. По
волокнам депрессорного нерва возбуждение достигает сосудодвигательного центра в
продолговатом мозгу, вызывая уменьшение частоты разрядов, следующих по
эфферентным волокнам к гладким мышцам стенок сосудов. Одновременно с этим
снижается частота сердечных сокращений. Итогом снижения тонуса сосудов и
появления брадикардии является понижение общего артериального давления.
|
|
|
Рис. 9.44 Влияние повышения давления в изолированном
каротидном синусе на артериальное давление собаки 1 —
давление в каротидном синусе; 2 — артериальное
давление; 3 — отметка времени 3 с. Цифрами справа обозначены величины
артериального давления, вверху — величины давления в каротидном синусе. |

Аортальная рефлексогенная зона является не только депрессорной,
как это вначале считали. При падении артериального давления частота импульсов в
депрессорном нерве уменьшается, что приводит к торможению центра блуждающего
нерва и активации симпатического. Разряды в последнем учащаются, что вызывает
сужение сосудов, стимуляцию деятельности сердца и в итоге — повышение общего
артериального давления. Эти два примера свидетельствуют о том, что
поддержание постоянства давления в аорте осуществляется авторегуляторными
механизмами, основанными на принципе отрицательной обратной связи.
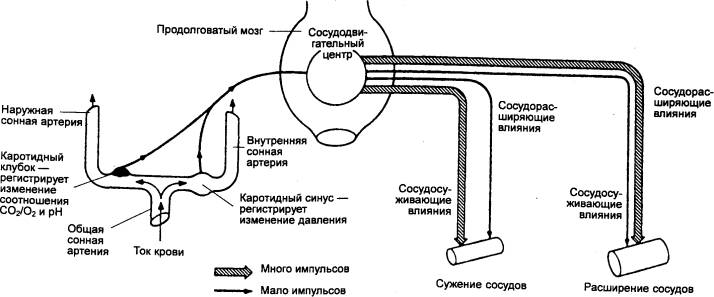
Область сонного (каротидного) синуса. Эта область — место разветвления общей сонной
артерии на внутреннюю и наружную (см. рис. 5.3) — вторая главная рефлексогенная
зона. Она была описана в 1923 г. Г. Герингом. Возбуждение барорецепторов зоны
каротидного синуса, возникающее в результате повышения давления в сонной
артерии, вызывает так называемый вазокардиальный рефлекс (рефлекс
Бейнбриджа), заключающийся в расширении сосудов и замедлении частоты
сердечных сокращений. Это, в свою очередь, приводит к общему снижению
артериального давления (рис. 9.44, 9.45).
Следовательно, чем выше давление в
дуге аорты и каротидном синусе, тем сильнее нервные влияния, ведущие к снижению
кровяного давления. Синокаротидная зона, подобно артериальной, может быть и
депрессорной, и прессорной. В случае падения давления в сонной артерии
интенсивность
|
|
|
Рис. 9.45 Взаимоотношения
между сонным (каротидным) клубком, каротидным синусом, сосудодвигательным
центром и общей системой кровообращения |
импульсации от барорецепторов уменьшается, что
сопровождается рефлекторным повышением тонуса мышц сосудистой стенки.
Соответственно повышается периферическое сопротивление сосудов и вследствие
этого нормализуется артериальное давление.
Сосуды легочного круга кровообращения. Как и в большом круге кровообращения, повышение
давления в легочной артерии при раздражении ее барорецепторов закономерно
приводит к брадикардии, гипотонии и расширению сосудов селезенки (рефлекс Парина),
что сопровождается снижением давления и, следовательно, устранением застоя
крови в легких.
В аортальной и каротидной рефлексогенных зонах наряду
с барорецепторами имеются еще и хеморецепторы, чувствительные к
изменению химического состава крови. Они открыты бельгийскими учеными Корнеем и
Жаном Хеймансами в 1926 г. Скопления сенсорных окончаний названы соответственно
аортальным и каротидным клубочками (гломусами). Хеморецепторы
дуги аорты представляют собой простое расположение клеток. В отличие от них
каротидный гломус имеет сложную капиллярную структуру с большим количеством
артериоло—венулярных анастомозов. Между капиллярами располагаются клетки двух
типов. Клетки I типа заполнены электронно—плотными везикулами, содержащими
биогенные амины. Клетки II типа не содержат аминов; своими отростками они
окружают клетки I типа. Собственно хеморецепторами считают клетки I типа и
свободные нервные окончания.
Хеморецепторы реагируют на изменение концентрации в
крови O2, C02, H+. Их возбуждение
может возникать также под влиянием некоторых неорганических и органических
веществ. Гипоксия, гиперкапния, которые сопровождаются изменением химического
состава крови, приводят к возникновению сердечно—сосудистых и дыхательных
рефлексов, которые направлены на нормализацию состава крови и поддержание
гомеостаза. Каротидные хеморецепторы в большей степени участвуют в регуляции
легочной вентиляции, аортальные — преимущественно в регуляции деятельности
сердечно—сосудистой системы. Как установлено В. Н. Черниговским, хеморецепторы
находятся также в сосудах сердца, селезенки, почек, костного мозга, органов
пищеварения и др. Их физиологическая роль состоит в восприятии концентрации
питательных веществ, гормонов, осмотического давления крови и передаче сигнала
об их изменении в ЦНС.
Механо— и хеморецепторы расположены также в стенках
венозного русла. Так, повышение давления в венах брюшной полости неизменно
сопровождается рефлекторным учащением и углублением дыхания, усилением
сердечного кровотока и присасывающего действия грудной клетки.
Рефлексы, возникающие с рецептивных зон сердечно—сосудистой
системы и определяющие регуляцию взаимоотношений в пределах именно этой
системы, носят название собственных (системных) рефлексов кровообращения (В.
Н. Черниговский). При небольших силах раздражения рецепторов они возникают без
сопровождения другими рефлексами (см. разд. 5.3.1). Так, незначительные
изменения давления в каротидных синусах влекут за собой прежде всего изменение
в системе кровообращения. Это будет собственный рефлекс кровообращения.
При увеличении силы раздражения в ответную реакцию помимо сердечно—сосудистой
системы вовлекается дыхание. Это будет уже сопряженный рефлекс. Пороги
раздражения для собственных рефлексов всегда ниже, чем для сопряженных. Переход
от собственных рефлексов к сопряженным совершается постепенно. Сопряженные
рефлексы рассматривают как определенную фазу в развитии цепи последовательных
рефлексов, вызываемых раздражением сосудистых рефлексогенных зон. Существование
сопряженных рефлексов дает возможность системе кровообращения быстро и
адекватно приспосабливаться к меняющимся условиям внутренней среды организма.
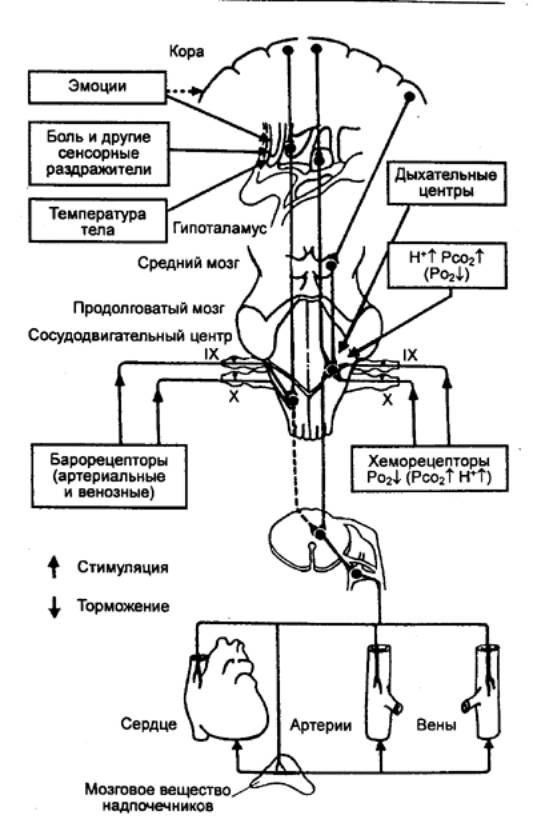
Центральное звено. Центральные механизмы, регулирующие поддержание артериального давления
на необходимом организму уровне, т. е. взаимодействие между величиной сердечного
выброса и тонуса сосудов, осуществляются за счет совокупности нервных структур,
которые принято называть сосудодвигательным (вазомоторным) центром. Это
понятие имеет собирательное функциональное значение, включающее различные
уровни центральной регуляции кровообращения с иерархической соподчиненностью
разных ее этажей. Структуры, относящиеся к вазомоторному центру, локализуются в
спинном, продолговатом мозгу, гипоталамусе, коре больших полушарий (рис. 9.46).
Спинальный уровень регуляции. Нервные клетки, аксоны которых образуют
сосудосуживающие волокна, располагаются в боковых рогах грудных и первых
поясничных сегментов спинного мозга. Эти клетки И. П. Павлов назвал спинальным
сосудосуживающим центром. Установлено, что при раздражении чувствительных
окончаний соматических нервов в спинном мозгу происходит передача возбуждения
на преганглионарные симпатические нейроны и таким образом осуществляется
замыкание рефлексов, регулирующих сосудистый тонус. Отделение спинного мозга от
продолговатого сопровождается падением величины кровяного давления, которое
вскоре восстанавливается до исходных показателей. Спинальные сосудосуживающие
нейроны поддерживают свой уровень возбудимости в основном импульсами от
вышерасположенных структур нервной
системы.
Бульварный уровень регуляции. Сосудодвигательный центр продолговатого мозга
является основным центром поддержания тонуса сосудов и рефлекторной регуляции
кровяного давления. Его локализация была установлена Ф. В. Овсянниковым (1871)
с помощью метода перерезок ствола мозга. У млекопитающих этот центр расположен
на дне IV желудочка головного мозга в его верхней части по обе стороны от
средней линии.
Сосудодвигательный центр подразделяется на
депрессорную, прессорную и кардиоингибирующую зоны. Депрессорная зона
способствует снижению артериального давления путем уменьшения активности
симпатических сосудосуживающих волокон, вызывая тем самым расширение сосудов и
падение периферического сопротивления, а также путем ослабления симпатической
стимуляции сердца, т. е. уменьшения сердечного выброса. Депрессорная зона
является местом переключения импульсов, поступающих сюда с
барорецепторов рефлексогенных зон, которые вызывают центральное торможение
тонических разрядов вазоконстрикторов. Кроме того, Депрессорная область
оказывает рефлекторное угнетение прессорной зоны и активирует парасимпатические
механизмы.
Прессорная зона оказывает прямо противоположное действие, повышая артериальное давление
через увеличение периферического сопротивления сосудов и сердечного выброса.
Взаимодействие депрессорных и прессорных структур сосудодвигательного центра
носит сложный синерго—антагонистический характер.
Кардиоингибирующее действие третьей зоны
опосредуется волокнами блуждающего нерва, идущими к сердцу. Его активность
приводит к уменьшению сердечного выброса и тем самым объединяется с активностью
депрессорной зоны в снижении артериального давления.
Деление сосудодвигательного центра на зоны довольно
условно, так как из—за взаимного перекрытия
зон определить границы невозможно. В одной и той же
|
|
|
Рис. 9.46 Важнейшие
«входы» и «выходы» сосудодвигательных центров
продолговатого мозга Эфферентные
волокна, усиливающие кровообращение, исходят преимущественно из
ипсилатеральных прессорных отделов, тогда как тормозные волокна от
депрессорных отделов пересекают продолговатый мозг и идут в вентралатеральных
участках спинного мозга на противоположной (контрлатеральной) стороне. |
зоне могут быть зарегистрированы и сосудорасширяющие,
и сосудосуживающие нейроны. Функциональное различие нейронов согласуется с
представлением о наличии депрессорной и прессорной зон.
Состояние тонического возбуждения
сосудодвигательного центра и, соответственно, уровень общего артериального
давления регулируются импульсами, идущими от сосудистых рефлексогенных зон
(рис. 9.47). Кроме того, этот центр входит в состав ретикулярной формации
продолговатого мозга, откуда также получает многочисленные коллатеральные
возбуждения от всех специфически проводящих путей.
Будучи автоматическим
саморегулирующимся центром, обеспечивающим основные элементарные функции
кровообращения (и в первую очередь оптимальный уровень давления в магистральных
сосудах), сосудодвигательный центр продолговатого мозга в реакциях целого
организма выступает в тесном взаимодействии с гипоталамусом, мозжечком,
базальными ядрами, корой головного мозга. Он осуществляет срочные ответы
кровеносной системы, связанные с усиленной мышечной работой, гипоксией,
гиперкапнией, ацидозом (Г. П. Конради). Высшая же форма регуляции
кровообращения, касающаяся энергетического обеспечения и интеграции
разнообразных сложных приспособительных актов и поведенческих реакций,
обеспечивается более высоким уровнем нервной системы.
|
|
|
Рис.
9.47 Нисходящие пути ствола мозга и
гипоталамуса, конвергирующие на преганглионарных нейронах промежуточной зоны пояснично—грудного
отдела спинного мозга (слева). Нервная регуляция артериального давления
(справа). «+», «—» — соответственно возбуждающие и тормозные эффекты
раздражения артериальных барорецепторов (Б) ЯОП —
ядро одиночного пути; ДЯ — дорсальное ядро блуждающего нерва; ДВЯ — двойное
ядро; ПВЯ — паравентрикулярное ядро гипоталамуса; РВЛ — рострально—вентралатеральная
область продолговатого мозга; IX — языкоглоточный нерв; Х — блуждающий нерв. |

Влияния самого сосудодвигательного центра осуществляются
через спинной мозг, ядра черепных нервов (VII, IX и Х пар), периферические
образования автономной нервной системы (рис. 9.48).
Гипоталамический уровень регуляции. Промежуточный мозг и особенно различные ядра
гипоталамической области рассматривают как сосредоточение высших подкорковых
центров, регулирующих функции разных отделов автономной нервной системы. С их
состоянием тесно связаны вазомоторные реакции гипоталамического происхождения,
играющие роль главным образом в осуществлении адаптивных реакций
кровообращения.
Интегративные центры гипоталамуса оказывают
нисходящее влияние на сердечно—сосудистый центр продолговатого мозга,
обеспечивая дифференцированный фазный и тонический контроль. Раздражение этого
отдела ведет к значительным изменениям кровяного давления. Лабораторные
животные v которых удалена вся кора мозга,
но сохранен промежуточный мозг, обнаруживают сложнокоординированные сосудистые
рефлексы в виде повышения кровяного давления и изменения частоты сердечных
сокращений.
В гипоталамусе, так же как в бульбарном
сосудодвигательном центре, различают депрессорные и прессорные
зоны (рис. 9.48). В целом это дает основание рассматривать гипоталамический
уровень как надстройку, выступающую в роли своеобразного дублера основного
бульбарного центра.
Корковый уровень регуляции. Влияние раздражения коры головного мозга на функции
кровообращения было обнаружено В. Я. Данилевским, Н. А. Миславским, В. М.
Бехтеревым, которые установили, что раздражение лобной и теменной областей
вследствие изменения сосудистого тонуса закономерно ведет к изменению кровяного
давления. Наиболее подробно эти реакции изучены с помощью методов условных
рефлексов. Так, сравнительно легко удается выработать сосудистую реакцию на
ранее индифферентный раздражитель, вызывая при этом ощущение жары, холода, боли
и т. д.
Определенные зоны коры головного мозга, как и
гипоталамус, оказывают нисходящее влияние на основной центр продолговатого
мозга. Эти влияния формируются в результате сопоставления информации, которая
поступила в высшие отделы нервной системы от различных рецептивных зон, с
предшествующим опытом организма. Они обеспечивают реализацию сердечно—сосудистого
компонента эмоций, мотиваций, поведенческих реакций.
Эфферентное звено. Эфферентная регуляция кровообращения реализуется через один и тот же
.аппарат, в основе которого лежат нервный и эндокринный механизмы.
Нервный механизм осуществляется, во—первых, при участии
преганглионарных симпатических нейронов, тела которых расположены в передних
рогах грудного и поясничного отделов спинного мозга, а также постганглионарных
нейронов, лежащих в пара— и превертебральных симпатических ганглиях. Вторым компонентом
являются преганглионарные парасимпатические нейроны ядра блуждающего нерва,
находящегося в продолговатом мозгу, и ядра тазового нерва, расположенного в
крестцовом отделе спинного мозга, и их постганлионарные нейроны. Третью часть
для полых висцеральных органов составляют эфферентные нейроны метасимпатической
нервной системы, локализующиеся в интрамуральных ганглиях их стенок. Названные
нейроны представляют собой общий конечный путь всех эфферентных и
центральных влияний, которые через адренергическое, холинергическое и другие
звенья регуляции действуют на сердце и сосуды.
Эндокринный отдел включает в себя
мозговой и корковый слои надпочечников, заднюю долю гипофиза,
юкстагломерулярный аппарат почек.
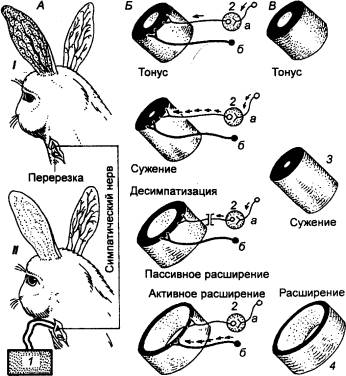
Впервые сосудосуживающее влияние симпатических
нервных волокон было показано отечественным физиологом А. П. Вальтером (1842)
на плавательной перепонке лягушки. Позже К. Бернар (1851) поставил специальные
опыты на ухе кролика. После перерезки симпатического нерва на шее кролика через
1—2 мин он наблюдал значительное расширение сосудов ушной раковины (рис. 9.49),
что проявлялось в покраснении кожи уха и повышении ее температуры. При
раздражении периферического конца этого перерезанного нерва кожа, покрасневшая
после перерезки симпатических волокон, становилась бледной и холодной на ощупь.
Это происходит в результате сужения просвета сосудов уха.
Тот же эффект характерен для сосудов органов брюшной
полости, почек, мозговых оболочек, кожи, однако в различных областях
сосудистого русла симпатический тонус проявляется не одинаково. Так, диаметр
сосудов кожи полностью зависит от симпатической иннервации, в то время как в
других органах эта зависимость проявляется в меньшей степени. Из этих опытов
следует, что симпатические волокна являются главными вазоконстрикторами, поддерживающими
тонус сосудов. Они постоянно находятся в состоянии тонического возбуждения,
поддерживая некоторую степень напряжения кольцевой сосудистой мышцы.
Особенно четко регуляторное значение симпатической
иннервации было установлено в экспериментах на симпатэктомированных кошках. В
случае незначительных кровопотерь у этих животных возникало резкое и стабильное
падение кровяного давления.
Симпатическая регуляция диаметра сосудов имеет
большое биологическое значение, так как, во—первых, она позволяет поддерживать
постоянным уровень кровяного давления, во—вторых, в зависимости от уровня метаболизма
регулировать кровоснабжение отдельных органов.
|
|
|
Рис.
9.48 Компоненты сосудодвигательного
центра А —
корковое представительство сосудодвигательного центра; Б — центры гипоталамуса: передний
гипоталамус (депрессорные зоны), задний гипоталамус (прессорные зоны); В — бульбарные центры
(сосудорасширяющий, сосудосуживающий); Г — центры спинного мозга (боковые
рога). 1 — кора больших
полушарий, 2 — ретикулярная
формация, 3 — сосудодвигательный центр, 4 — спинной мозг, 5 — симпатический ганглий, 6 — кровеносные сосуды, 7 —
продолговатый мозг, 8 —
гипофиз, 9 — гипоталамус. |
|
|
|
Рис.
9.49 Механизмы сосудодвигательных
реакций А — влияние
симпатического нерва (опыт К. Бернара): I — результат десимпатизации, II — результат раздражения периферического
конца перерезанного симпатического нерва; Б — нервная регуляция просвета сосуда: а —
сосудосуживающие симпатические нервы (адренергические), б — сосудорасширяющие нервы; В — гуморальная регуляция просвета
мелких сосудов; 1 —
стимулятор, 2 — симпатический
ганглий, 3 — норадреналин, ангиотензин, вазопрессин и др., 4 — СО2, молочная
кислота, гистамин, брадикинин и др. |

Степень сокращения сосудистых гладких мышц зависит от
частоты разрядов в сосудодвигательных волокнах. Следовательно, возрастание
числа импульсов приводит к вазоконстрикции. Однако, если уменьшение числа
импульсов идет от некоторого уровня, наступает дилатация. Дилатация не
является беспредельной, она ограничена базальным тонусом сосудов. После
симпатэктомии в денервированном участке также наблюдается вазодилатация, и в
этом случае диаметр сосудов целиком определяется базальным тонусом.
Помимо вазоконстрикторных волокон существуют
специальные волокна возбуждение которых сопровождается вазодилатацией. Эти
волокна были идентифицированы М. Шиффом и позже К. Бернаром при раздражении барабанной
струны — веточки язычного нерва, идущей к поднижнечелюстной слюнной железе.
Раздражение веточки вызывает сильное расширение сосудов железы.
Аналогичный эффект возникает при стимуляции других
нервов — языкоглоточного, верхнегортанного, тазового. Все они являются
парасимпатическими. Однако это вовсе не означает, что все вазодилататоры
относятся к парасимпатической нервной системе и что все парасимпатические
волокна расширяют сосуды. Например, волокна парасимпатического блуждающего
нерва сужают сосуды сердца. К закономерному расширению сосудов сердца и
скелетных мышц может привести возбуждение симпатических волокон. Это свидетельствует
о том, что указанные волокна относятся к холинергическим волокнам
симпатического происхождения. Имеются сосудорасширяющие волокна и в
соматических нервах, таких как седалищный, однако из—за присутствия в этом
стволе вазоконстрикторов их трудно обнаружить. Нельзя не учитывать, что сосуды
способны к расширению и без участия дилататоров, а, как было показано выше,
просто в результате снижения вазоконстрикторной активности.
Иннервация вен соответствует иннервации артерий, хотя
в целом плотность иннервации вен значительно меньше. Нервные окончания
эфферентных волокон точно прослежены до прекапиллярных сфинктеров, где они
оканчиваются на гладкомышечных клетках. Сфинктеры способны активно отвечать на
проходящие импульсы.
Основным механизмом нервной регуляции капилляров является эфферентная иннервация бессинаптического
типа посредством свободной диффузии медиаторов в направлении стенки сосуда. В
зависимости от расстояния, которое проходит медиатор, нервные влияния на
капилляры могут быть быстрыми и непосредственными или замедленными и
опосредованными. Распространяясь во все стороны от терминали, медиатор влияет
на все части функционального элемента в пределах целого микроучастка.
Эндокринная регуляция. Главную роль в эндокринной регуляции сосудистого
русла играют гормоны— мозгового и коркового слоев надпочечников, задней
доли гипофиза и юкстагломерулярного аппарата почек.
Адреналин
обладает резким сосудистым действием. На артерии и артериолы кожи, органов
пищеварения, почек и легких он оказывает сосудосуживающее влияние, на
сосуды скелетных мышц, гладкой мускулатуры бронхов — расширяющее,
содействуя тем самым перераспределению крови в организме. При физическом
напряжении, эмоциональном возбуждении он способствует увеличению кровотока
через скелетные мышцы, мозг, сердце.
Адреналин, как и норадреналин, выделяющийся в
постганглионарных симпатических окончаниях, иннервирующих кровеносные сосуды,
активирует аденилатциклазу, которая находится на внешней мембране мышечных
клеток. Через образовавшийся 3,5—АМФ он вызывает сужение сосудов.
Влияние адреналина и норадреналина на сосудистую
стенку определяется существованием разных типов адренорецепторов — α и
β, представляющих собой участки гладкомышечных клеток с особой химической
чувствительностью. В сосудах обычно имеются оба типа рецепторов. Взаимодействие
медиатора с α—адренорецептором ведет к сокращению стенки сосуда, с (β—рецептором
— к расслаблению. Норадреналин взаимодействует в основном с альфа—адренорецепторами,
адреналин — с α— и (β—рецепторами. По мнению У. Кеннона, адреналин —
это «аварийный гормон», осуществляющий в трудных, иногда экстремальных условиях
мобилизацию функций и сил организма.
Альдостерон — другое необходимое звено регуляции кровообращения железами
надпочечников. Он вырабатывается в их корковом слое. Альдостерон обладает
необычайно высокой способностью усиливать обратное всасывание натрия в почках,
слюнных железах, пищеварительной системе, изменяя таким образом
чувствительность стенок сосудов к влиянию адреналина и норадреналина.
Вазопрессин — гормон задней доли гипофиза. Он вызывает сужение артерий и артериол
органов брюшной полости и легких. Однако, как и под влиянием адреналина, сосуды
мозга и сердца реагируют на этот гормон расширением, что способствует улучшению
питания и мозговой ткани, и сердечной мышцы.
Клетки юкстагломерулярного аппарата почек продуцируют
фермент ренин. Он обнаружен у некоторых видов костистых рыб, амфибий,
рептилий. У млекопитающих в цепи ферментативных реакций с участием глобулинов
крови он превращается в ангиотензин II. Ангиотензин II обладает мощным
вазоконстрикторным действием, значительно превосходящим по силе норадреналин,
но в отличие от последнего не вызывает выброса крови из депо. Это объясняется
наличием чувствительных к ангиотензину рецепторов только в прекапиллярных
артериолах, которые расположены в организме неравномерно. Поэтому его действие
на сосуды различных областей не одинаково. Системный прессорный эффект
сопровождается уменьшением кровотока в почках, кишке, коже и увеличением его в
мозгу, сердце и надпочечниках. Изменения кровотока в мышце незначительны.
Большие дозы ангиотензина могут вызвать сужение сосудов сердца и мозга. Ренин и
ангиотензин представляют собой ренин—ангиотензиновую систему.
Помимо прямого действия на сосудистую систему
ангиотензин оказывает влияние и опосредованно через автономную нервную систему
и эндокринные железы. Он увеличивает секрецию альдостерона, адреналина и
норадреналина, усиливает вазоконстрикторные симпатические эффекты.
Способностью расширять сосуды обладают биологически активные
вещества и местные гормоны, такие как гистамин, серотонин, брадикинин,
простагландины.
Гистамин
содержится главным образом в тучных клетках и базофильных гранулоцитах. Он
расширяет сосуды сердца, печени, кишки, увеличивает наполнение капилляров, повышает
проницаемость их эндотелия, а также уменьшает объем циркулирующей крови.
Действием гистамина объясняется реакция покраснения кожи. При введении его в
организм возникает резкое снижение артериального давления, сходное с состоянием
организма при обильном кровотечении или возникновении гистаминового шока.
Серотонин
найден у некоторых беспозвоночных и почти у всех позвоночных животных. Главным
местом его образования являются энтерохромаффинные клетки кишки. В зависимости
от вида животного и условий эксперимента изменения артериального давления могут
иметь фазный характер: гипотензия сменяется гипертензией, после которой вновь
развивается гипотензия. Это объясняется тем, что на тонус сосудов серотонин
оказывает прямое и рефлекторное действие. Прямое действие на гладкую
мускулатуру приводит к спазму сосудов и повышению артериального давления,
воздействие на рефлексогенные зоны вызывает гипотензию.
Брадикинин
образуется в плазме крови, но особенно много его в поднижнечелюстной слюнной и
поджелудочной железах. Являясь регуляторным пептидом он расширяет сосуды кожи,
скелетных мышц, мозговые и коронарные сосуды.
Простагландины представляют большую группу биологически активных веществ. Они являются
производными ненасыщенных жирных кислот. Простагландины образуются практически
во всех органах и тканях, однако термин для их обозначения связан с
предстательной железой, из которой они были впервые выделены. Биологическое
действие простагландинов чрезвычайно многообразно. Один из их эффектов
проявляется в выраженном действии на тонус гладкой мускулатуры сосудов, причем
влияние разных типов простагландинов часто диаметрально противоположно. Одни
Простагландины сокращают стенки кровеносных сосудов и повышают артериальное
давление, другие — оказывают сосудорасширяющее действие, сопровождающееся
гипотензивным эффектом.
В нервной и эндокринной регуляции различают
гемодинамические механизмы кратковременного действия, промежуточные и
длительного действия.
К механизмам кратковременного действия относят
циркуляторные реакции нервного происхождения — барорецепторные,
хеморецепторные, рефлекс на ишемию ЦНС. Их развитие происходит, в течение
нескольких секунд. Промежуточные (по времени) механизмы охватывают
изменения транскапиллярного обмена, расслабление напряженной стенки сосуда, реакцию
ренин—ангиотензиновой системы. Для включения этих механизмов требуются минуты,
а для максимального развития — часы. Регуляторные механизмы длительного действия
влияют на соотношение между внутрисосудистым объемом крови и емкостью сосудов.
Это осуществляется посредством транскапиллярного обмена жидкости. В этом
процессе участвуют почечная регуляция объема жидкости вазопрессин и
альдостерон.