8.4.3.
Основные свойства некоторых цитокинов
ИЛ1. Впервые
этот плейотропный медиатор был обнаружен в перитонеальном (брюшинном) экссудате
в острую фазу воспаления. После введения его животным наблюдается повышение
температуры тела. В связи с этим ИЛ1 получил название «эндогенный пироген».
Позднее эндогенный пироген был очищен. Дальнейшие исследования позволили
обнаружить новые эффекты влияния эндогенного пирогена: уменьшение уровня цинка
и железа в плазме крови, нейтрофилию, стимуляцию синтеза амилоидного белка А в
печени, колониестимулирующую активность. Эндогенный пироген был описан как
«лимфоцит—активирующий фактор», являющийся костимулятором Т—лимфоцитов . Стандартным
биологическим тестом для обнаружения ИЛ1 служит синергичная стимуляция
пролиферации мышиных тимоцитов в комбинации с митогенными лектинами
растительного происхождения, такими как конканавалин А или фитогемагглютинин.
На основании исследований, посвященных продукции ИЛ1, главными его
источниками считались моноциты и макрофаги. Однако позднее было обнаружено, что
ИЛ1 продуцируется и другими клетками, имеющими с макрофагами общее
происхождение: звездчатые ретикулоэндотелиоциты (клетки Купфера) в печени,
белые отростчатые эпидермоциты (клетки Лангерганса) в эпидермисе, клетки
микроглии и др. К числу продуцентов этого медиатора относятся фибробласты. Т— и
В—лимфоциты, натуральные киллеры, нейтрофилы и эндотелиоциты. Установлено, что
нейроны и астроциты способны стимулировать и высвобождать ИЛ1. Продукция ИЛ1
осуществляется периферическими симпатическими нейронами и хромаффинными
клетками мозгового вещества надпочечников и пищеварительного тракта.
ИЛ1 существует в виде двух различных полипептидных форм ИЛ1 а и ИЛ1β, синтез которых кодируется двумя генами. Обе эти формы образуются из соответствующих молекул—предшественников, имеющих одинаковую молекулярную массу (31 кДа). Предшественник ИЛ1α (проИЛ1α) биологически активен и способен соединяться с рецептором в форме димера. ИЛ1β приобретает способность связываться с рецептором только после ферментативного расщепления, в результате которого образуется конечный продукт с молекулярной массой 17,5 кДа. Этот процесс катализируется ИЛ1—β—конвертирующим энзимом.
Преобладающей формой ИЛ1 является ИЛ1β: количество соответствующей
мРНК, обнаруживаемое в активированных клетках, в 1050 раз превышает количество
мРНК для ИЛla. В
неактивированных клетках, как правило, не находят значительных количеств мРНК,
кодирующих синтез ИЛ1. Резкое увеличение продукции цитокина происходит в ответ
на инфекцию, действие микробных токсинов, медиаторов воспаления, продуктов
активированных лимфоцитов и комплемента.
Несмотря на структурные различия ИЛla и ИЛ1β, разную продолжительность их жизни (ИЛlα —
15 ч, ИЛ1β — 2,5 ч), две формы цитокина связываются с одними и теми же
рецепторами. Обнаружено два типа рецепторов для ИЛ1: на Т—лимфоцитах и
фибробластах с молекулярной массой 80 кДа и на В—лимфоцитах с молекулярной
массой 68 кДа. Оба типа рецепторов принадлежат к семейству иммуноглобулинов, их
внеклеточные домены содержат три домена гомологичных иммуноглобулинов.
ИЛ1 играет одну из центральных ролей в воспалительной реакции, в ответе
на бактериальную инфекцию и тканевые повреждения. Он проявляет свойства
нейроэндокринного гормона, стимулируя продукцию кортикотропина, простагландинов
и целого ряда других гормонов. Сообщалось, что ИЛ1 является митогеном для
астроцитов. Этот цитокин стимулирует продукцию ИЛ2 Т—лимфоцитами, повышает
экспрессию рецептора для ИЛ2. ИЛ1 усиливает пролиферацию В—лимфоцитов, секрецию
антител и экспрессию мембранного иммуноглобулинового рецептора. Под влиянием
ИЛ1 возрастает секреция гепатоцитами сывороточных амилоидов А и Р, С—реактивного
белка, α1—антитрипсина и церулоплазмина. Связывание ИЛ1 с рецепторами
сопровождается целым рядом других биологических эффектов: повышением
температуры тела, нарушением сна, головной болью, нарушениями деятельности
пищеварительной системы. Модуляция под действием ИЛ1 тех или иных процессов
осуществляется путем повышения или понижения уровня экспрессии других
рецепторов. ИЛ1 усиливает связывание опиатов рецепторами, повышает экспрессию
рецепторов для TFGJ3
(трансформирующего фактора роста), понижает экспрессию ИЛ1—рецепторов I типа,
уменьшает число рецепторов ФНО, снижает количество мРНК, кодирующей синтез
рецепторов ИЛ6, ингибирует образование рецепторов для лютеинизирующего гормона.
ИЛ2. Этот цитокин с молекулярной массой 15 кДа играет
исключительно важную роль в реализации механизмов иммунного ответа. У различных
видов животных и человека продукция ИЛ2 осуществляется лимфоцитами
периферической крови, костного мозга, лимфатических узлов, перитонеального
экссудата, тимуса, селезенки, миндалин и грудного протока, стимулированными к
пролиферации антигенами или лектинами. При этом продуцентами ИЛ2 являются T—хелперы
1. Покоящиеся лимфоциты не содержат и спонтанно не продуцируют ИЛ2. Обычный
физиологический путь активации лимфоцитов, обусловленный взаимодействием
специфического антигена и ТКР, не является единственным путем стимуляции Т—лимфоцитов
. T—лимфоциты могут быть стимулированы к продукции ИЛ2 с помощью ряда
митогенных лектинов (фитогемагглютинин, конканавалин А), антител к поверхностным
маркерным молекулам и целого ряда других биологически активных веществ.
Сообщалось о значительном усилении продукции ИЛ2 лимфоцитами и гормональными
факторами тимуса.
Помимо участия ИЛ2 в дифференцировке и пролиферации Т—лимфоцитов , этот
лимфокин вносит важный вклад в реализацию механизмов противоопухолевой
защиты. Он повышает литическую активность нормальных киллеров, а также
индуцирует клетки системы лимфокин—активированных киллеров. Кроме того, ИЛ2
вызывает пролиферацию ЦТЛ. Под действием ИЛ2 усиливается секреция ИФНγ Т—лимфоцитами.
Таким образом, ИЛ2 является ключевым фактором в развитии иммунологических
реакций. Действие ИЛ2 приводит к развитию плейотропного эффекта с участием
нескольких субпопуляций иммунной системы (рис. 8.53).
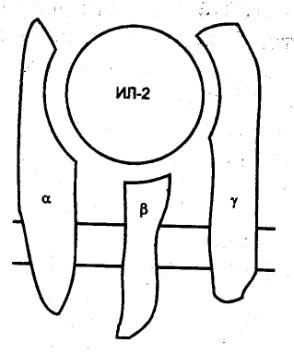
Рецептор для ИЛ2 состоит из трех полипептидных цепей, которые могут быть
экспрессированы вместе и по отдельности (рис. 8.54). В зависимости от того, из
скольких цепей будет составлен рецептор, изменяется его аффинность. Так, при
экспрессии одной α—цепи (молекулярная масса 55 кДа) рецептор обладает
наиболее низкой аффинностью с константой диссоциации (Kd) около 10 нМ. Такой рецептор очень небольшую
цитоплазматическую часть и после связывания ИЛ2 не способен проводить сигнал
внутрь клетки.
ИЛ3. Этот цитокин относится к семейству гемопоэтических ростовых
факторов (молекулярная масса 1528 кДа), является колониестимулирующим фактором
гранулоцитов—макрофагов. ИЛ3 вместе с эритропоэтином поддерживает рост и
дифференцировку клеток эритроидного ряда. Он способен регулировать раннюю
стадию дифференцировки В—лимфоцитов, поддерживает рост пре—В—клеток, а также
усиливает секрецию IgG.
|
|
|
Рис. 8.53 Схема биологических эффектов ИЛ2 Аг—
антиген; Мф — макрофаг; Тл — Т—лимфоцит; Вл — В—лимфоцит; — натуральные
киллеры; ЛАК — лимфокин—активированные киллеры; ФНО— фактор
некроза опухолей, Iа —
молекулы МНС II класса. |
|
|
|
Рис. 8.54 Строение высокоаффинного рецептора для ИЛ2 Для проведения сигнала требуется гетеродимеризация
цитоплазматических доменов β— и γ—цепеи. |

По химической природе ИЛ3 —
гликопротеин, углеводный компонент содержит сиаловые кислоты. Основным
источником ИЛ3 являются T—лимфоциты с фенотипическими характеристиками хелперов,
активированные фитогемагглютинином или антигеном.
ИЛ3 — один из основных факторов, поддерживающих рост и жизнеспособность
предшественников тучных клеток, играющих важную роль в нормальной
жизнедеятельности организма и при патологии. ИЛ3 мыши и человека выполняют
функцию фактора, активирующего клетки к продукции гистамина. Не исключено, что
вызываемая им активация пролиферации полипотентных клеток костного мозга
связана со способностью ИЛ3 стимулировать в клетках синтез гистамина. Благодаря
активации костномозгового кроветворения ИЛ3 обладает радиопротекторными
свойствами. Он потенцирует цитотоксическую активность макрофагов, вызванную
липополисахаридом.
ИЛ4. Этот лимфокин (молекулярная масса 1520 кДа) продуцируется Т—хелперами
2 и является фактором дифференцировки для Т— и В—лимфоцитов. Он служит
кофактором пролиферации покоящихся В—лимфоцитов и индуцирует продукцию
отдельных изотипов иммуноглобулинов, в частности, IgE и IgG
ИЛ4 служит ростовым фактором для части Т—лимфоцитов , стимулирует пролиферацию
костномозговых клеток—предшественников гемопоэза и тучных клеток. Известна
способность ИЛ4 генерировать активность ЛАК и усиливать противоопухолевую
активность макрофагов. Помимо Т—лимфоцитов , ИЛ4 может также продуцироваться
тучными клетками и точно не идентифицированными клетками стромы костного мозга.
Важной
особенностью ИЛ4, отличающей его от многих других цитокинов является строгая
видовая специфичность биологического действия. ИЛ4 человека оказывает
биологическое действие и связывается только с клетками человека и обезьян.
Мышиный ИЛ4 активен только в отношении различных линий мышей, но не действует
на клетки приматов.
ИЛ4 играет роль регулятора
иммунологических функций, способствующего достижению оптимального уровня
иммунного ответа. Однако его гиперпродукция или дисбаланс регуляторных влияний
со стороны других цитокинов могут проявляться в форме повышенной
иммунологической реактивности.
ИЛ5. Этот
димерный белок с молекулярной массой 5060 кДа продуцируется Т—хелперами 2, т.
е. теми же клетками, что и ИЛ4. Синтез ИЛ5 Т—хелперными лимфоцитами происходит
только в ответ на антигены или другие индукторы.
Влияние ИЛ5 на эозинофилы
заключается в усилении функциональной активности зрелых клеток, поддержании их
жизнеспособности, продукции супероксидного аниона и стимуляции хемотаксиса
эозинофилов, а также избирательной стимуляции пролиферации и дифференцировки
предшественников эозинофилов в костном мозгу. В последнем ИЛ5 синергичен с ИЛ1
и ИЛ3. Вероятно, ИЛ5 является цитокином, ответственным за селективную активацию
эозинофилов при аллергических заболеваниях.
ИЛ6. Этот
цитокин представляет собой белок с молекулярной массой 1934 кДа. Он
секретируется многими типами клеток, участвующих прямо или косвенно в
осуществлении воспалительной реакции и иммунного ответа (покоящиеся T—лимфоциты;
T—лимфоциты, трансформированные вирусом; фибробласты; моноциты; макрофаги;
эндотелиоциты; кератиноциты; клетки многих опухолей). Действие ИЛ6 может
осуществляться местно в очаге воспаления и системно при его попадании в циркуляцию.
Местное действие ИЛ6 на
иммунокомпетентные клетки заключается в стимуляции пролиферации T—хелперов 1 и
2, усилении продукции ИЛ2, индукции ЦТЛ, а также в ускорении дифференцировки В—лимфоцитов
в антителопродуценты. ИЛ6 увеличивает экспрессию молекул МНС класса I, но не
класса II.
Системное действие ИЛ6,
характеризующее его как типичный медиатор воспалительной реакции,
проявляется при распространении очага инфекции и попадании ИЛб в циркуляцию.
Одним из важных звеньев системного действия ИЛ6 служит его стимулирующее
влияние на пролиферацию костномозговых клеток—предшественников гранулоцитов и
макрофагов, приводящее к увеличению выхода зрелых форм этих клеток для
восполнения утраченных в ходе воспалительной реакции фагоцитов.
ИЛ6 вызывает индукцию белков острой
фазы воспаления гепатоцитами. Эта особенность цитокина также является
проявлением его системного действия. ИЛб, так же как ИЛ1 и ФНО, индуцирует
лихорадку, являясь еще одним эндогенным пирогеном. Он оказывает прямое действие
на клетки мозга, индуцируя продукцию АКТГ.
ИЛ7. Этот
лимфокин является полипептидом с молекулярной массой 14,9 кДа. Биологические
свойства ИЛ7 изучены недостаточно. Полагают, что этот цитокин выступает одним
из факторов, продуцируемых клетками стромы костного мозга, а возможно, и
тимуса, которые стимулируют рост пре—В— и пре—Т—лимфоцитов .
ИЛ8.
Представляет собой олигопептид с молекулярной массой 8,8 к Да. Его
изоэлектрическая точка находится в области 8,08,5. Как и все интерлейкины, он
является индуцибельным белком. ИЛ8 продуцируется макрофагами, лимфоцитами,
эпителиальными клетками почки, фибробластами, эндотелиоцитами. Рецепторы для
ИЛ8 обнаружены на нейтрофилах. ИЛ8 является мощным индуктором острой
воспалительной реакции, вызывая миграцию нейтрофилов в очаг проникновения патогенного
фактора. Помимо основной функции, ИЛ8 способствует усилению адгезивных свойств
нейтрофилов, генерации активных форм кислорода, активации пентозофосфатного
шунта, подъему концентрации Са2+, изменяет форму нейтрофилов,
способствует экзоцитозу.
Фактор
некроза опухолей (ФНО). Был впервые получен в результате введения мышам
бактериальных эндотоксинов. Обладает цитотоксической активностью по отношению к
опухолевым клеткам. ФНО представляет собой полипептид с молекулярной массой
около 17 кДа. Основными продуцентами этого цитокина являются моноциты
(макрофаги) и лимфоциты. Под воздействием бактериальных токсинов клетки
моноцитарно—макрофагальной системы активируются и начинают синтезировать ФНО,
ИЛ—1, ИЛ—6 и вещества прокоагулянтного действия.
Многие функции ФНО—α и
ФНО—β (лимфотоксин) идентичны функциям ИЛ—1. При совместном использовании
этих цитокинов часто наблюдается потенцирование действия или синергизм между
молекулами. ФНОα и ИЛ1 действуют синергично на фибробласты, усиливая
продукцию ПГЕ2. Синергичное влияние выявлено при радиопротекторном
эффекте. Полагают, что такой синергизм является скорее всего результатом
воздействия вторичных мессенджерных молекул, так как ИЛ1 уменьшает число
рецепторов для ФНОа.
ФНОα и
ФНОβ способны тормозить клеточную пролиферацию, дифференцировку и функции
клеток почти всех типов. Они служат медиаторами цитотоксичности. В результате
их высвобождения повышается проницаемость капилляров, повреждается эндотелий
сосудов, возникает внутрисосудистый тромбоз. В настоящее время признано, что
ФНО играет ведущую роль в патогенезе септического шока.
ИЛ10. Этот
лимфокин (1721 кДа), продуцируемый Т—хелперами 2, может рассматриваться как
антагонист ряда цитокинов. ИЛ10 подавляет продукцию ИФНγ Т—хелперами 1. Оh тормозит
пролиферативный ответ T—клеток на антигены и митогены, а также подавляет
секрецию ИЛ1β, ФНО и ИЛ6. В то же время ИЛ10 стимулирует секрецию
иммуноглобулинов В—лимфоцитами.
ИЛ12. Лимфокин
человека состоит из двух субъединиц с молекулярной массой 40 и 50 кДа, удерживаемых
дисульфидной связью. Субъединица р40 гомологична рецептору для ИЛ6, а
субъединица р50 имеет частичную гомологию с ИЛ6 и гранулоцитарным
колониестимулирующим фактором. ИЛ12 повышает литическую активность клеток
системы ЛАК, а также специфическую цитотоксичность ЦТЛ. ИЛ12 рассматривают как
ростовой фактор при активации Т—лимфоцитов
и натуральных киллеров.