Механизмы действия гормонов
наиболее удобно рассмотреть на примере трех эндокринных клеток, секретирующих
гормоны в межклеточное пространство в непосредственной близости от капилляров
(рис. 6.31).
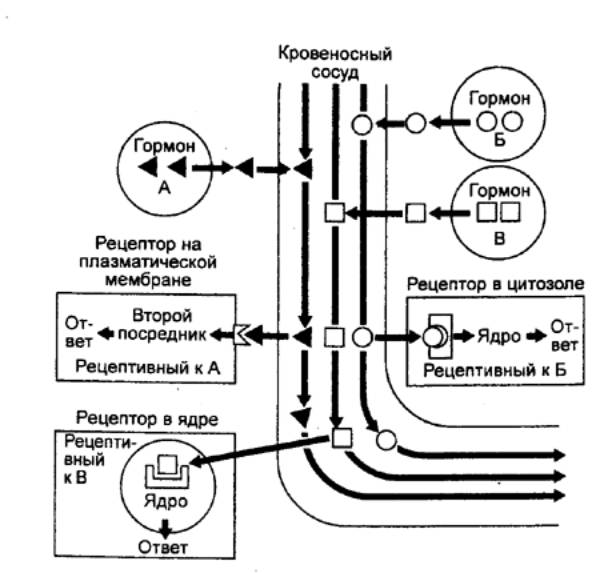
Рецептором называют молекулу (или их комплекс),
способную к концентрационно зависимому связыванию лиганда (гормон, медиатор) и
передаче сигнала об этом событии с его усилением и изменением (трансдукцией) к
внутриклеточным структурам. Взаимодействие гормона с внутриклеточным рецептором
представляется следующим образом (рис. 6.31). Гормон попадает в клетку через
плазматическую мембрану и взаимодействует с рецептором. Гормон—рецепторный
комплекс переносится в ядро и действует на синтез ДНК или же изменяет скорость
транскрипции и синтез информационной (матричной) РНК (мРНК). Увеличение или
уменьшение количества мРНК влияет на синтез белка в процессе трансляции, что
приводит к изменению функциональной активности клетки.
Активность рецептора
характеризуется сродством (специфичностью связывания) к лиганду и способностью
воздействовать на эффекторные механизмы клетки прямо или через молекулы—посредники.
Большинство известных рецепторов является белками, фосфо— или гликопротеинами,
имеющими один участок связывания определенного лиганда.
Для каждого из гормонов
известны несколько типов рецепторов, локализованных преимущественно в клетках
разных органов—мишеней. Это во многих случаях связано со сменой механизма
трансдукции гормонального сигнала или системы инактивации рецептора. Например,
глутамат, наиболее распространенный возбуждающий нейротрансмиттер, через метил—П—аспартат
(NMDA)—рецептор, ионный канал, вызывает быстрые реакции нейронов.
Другой тип рецепторов глутамата, метаботропический (mGluR), сопряжен с ГТФ—связывающими
белками и опосредует более медленные реакции через системы внутриклеточных
посредников.
Эффекты лиганд—рецепторного взаимодействия в
значительной степени определены локализацией рецептора, который может
находиться в плазмалемме и внутриклеточных мембранах, цитозоле и нуклеоплазме
(рис. 6.32). При
|
|
|
Рис. 6.31 Механизм
действия гормонов |
|
|
|
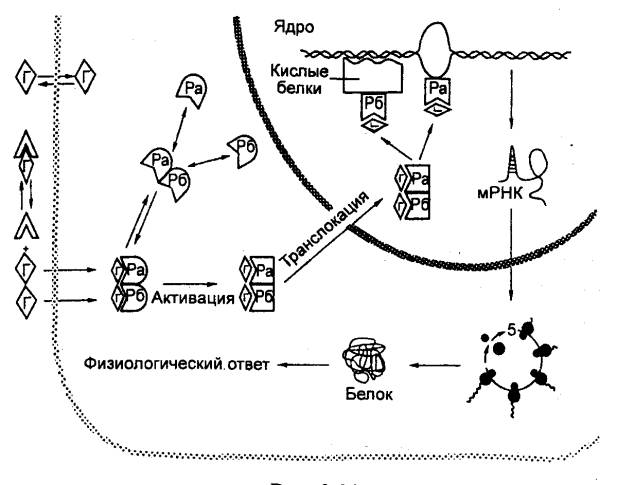
Рис. 6.32 Механизм
действия стероидных гормонов Ра и Рб — две субъединицы рецептора; Г —
гормон. |
эндоцитозе лиганд—рецепторного
комплекса гормон может попадать в клетку и эффекты мембранного уровня
дополняются внутриклеточными.
Известно,
что одновременно на клетку могут действовать многие вещества, взаимодействуя с
разными рецепторами и определяя таким образом вероятностный характер итоговой
реакции. Однако существует множество факторов, увеличивающих вероятность
определенного функционально значимого ответа клетки через выделение и усиление
наиболее значимого сигнала, регламентацию длительности его воздействия и определение
последовательности событий на мембране и в клетке. К таким факторам,
действующим на уровне мембраны, относят локальные значения: потенциала
мембраны, градиентов рН, осмотического давления и ионов.
Избирательному
усилению действия одного из сигналов может способствовать и увеличение числа
определенных рецепторов под влиянием повторяющихся воздействий данного гормона
либо вследствие предварительного воздействия другого. Например, эстрадиол
влияет на экспрессию генов и синтез рецепторов к окситоцину и базальному
фактору роста фибробластов, что затем приводит к повышению эффективности
действия этих гормонов. Увеличение числа рецепторов приводит к образованию их
групп (кластеров) благодаря латеральной диффузии молекул в плазмалемме,
действию белков кластрирования, рецептор—зависимому фосфорилированию белковых
факторов кластеризации. Для рецепторов факторов роста в таких кластерах
показана возможность взаимного фосфорилирования. Сходная кластеризация описана
и для хемочувствительных ионных каналов, активируемых гормонами. По механизмам,
опосредующим эффекты связывания лиганда и рецептора, выделены три основные типа
рецепторов, локализованных в клеточной мембране.
Рецепторы
I типа
представлены хемочувствительными ионными каналами, одна из субъединиц которого
обладает способностью связывать лиганд. Так, для медиатора нервной системы и
парагормона многих эндокринных желез ГАМК известен рецептор А—типа, α—субъединица
которого имеет консервативный трансмембранный домен М II, содержащий ряд
остатков аминокислот Thr и Ser и составляющий основу Cl——канала или водной поры. Его
другие субъединицы выполняют рецептивные функции. В результате лиганд—рецепторного
взаимодействия ионы хлора входят в клетку и гиперполяризуют мембрану, а
сопряженный транспорт воды участвует в регуляции объема и осмотического
давления в клетке.
Рецепторы
II типа —
интегральные белки с семью трансмембранными доменами. Участок внутриклеточного
С—конца молекулы рецептора и одна из внутриклеточных петель (чаще третья)
сопряжены с одной из трех (а) субъединиц ГТФ/ГДФ—связывающего (G)
белка
клеточной мембраны. Связывание гормона приводит к рецептор—зависимой активации
внутриклеточных факторов, обусловливающих замену ГДФ на ГТФ в неактивном
комплексе ГДФ/α—субъединица. При этом две другие субъединицы G—белка
((β и γ) диссоциируют от комплекса и могут воздействовать на ионные
каналы мембраны. Другая группа внутриклеточных факторов (ГТФаз—активирующие
белки, GAP) активирует энзиматические
свойства α—субъединицы, и она как ГТФаза отщепляет фосфатную группу от
ГТФ. Выделяемая при этом энергия идет на фосфорилирование энзимов, а также
белков цитоскелета и других белков.
Разные
изоформы α—субъединицы гетеротримерных G—белков клеточных мембран
воздействуют на определенные энзимы, запуская соответствующие каскады
посредников. Таким образом осуществляется передача, усиление и трансформация
сигнала, кодируемого гормоном или другим лигандом.
|
|
|
Рис. 6.33 Механизм
проведения гормонального сигнала в клетке при участии вторичных посредников Gs
— Gs—белок, G1 — Gi—белок, Gq — Gq—белок; ИФЗ — инозитолтрифосфат, КМ —
кальмодулин, Ф — фосфат; α1, α2, β — субтипы рецепторов. |
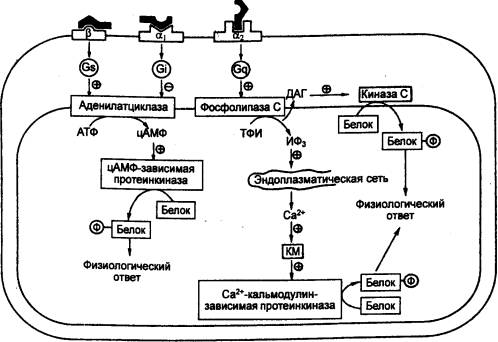
В аденилатциклазной системе рецептор—зависимая
активация Gs—белка приводит к активации энзима мембраны аденилатциклазы,
которая конвертирует АТФ в цАМФ (рис. 6.33). Последняя фосфорилирует
цитоплазматические белки. Один из них — цАМФ—зависимая протеинкиназа А
(РКА), каскадно—фосфорилирующая по остаткам Ser и Thr молекулы других белков,
выполняющих разнообразные функции.
Рецептор—зависимая активация Gi—белка (его G2i
изоформы) приводит к подавлению активности аденилатциклазы. Фосфорилирование
α—субъединицы G2i по Ser144 (серин—треониновой
протеинкиназой С, активированной, например, действием гормона вазопрессина)
блокирует тормозные эффекты Gi в гепатоцитах и
тромбоцитах. Аденилатциклазную систему вторичных посредников могут подавлять
также фосфодиэстераза, превращающая цАМФ в быстро гидролизируемый мономер,
и лиганды, активирующие гуанилатциклазную систему.
В гуанилатциклазной системе соответствующий энзим,
конвертирующий ГТФ в цГМФ, представлен двумя пулами: мембранной и цитоплазматической
гуанилатциклазами. В плазмалемме энзим может быть компонентом рецептора
(например, рецептора атриального натрийуретического гормона) или сопряжен с G—белком.
Цитозольная гуанилатциклаза также обладает рецепторными функциями,
акцептируя такое вещество, как оксид азота (NO). В обоих случаях лиганд—рецепторное
взаимодействие приводит к активации энзима и увеличению внутриклеточной
концентрации цГМФ. Последняя, как и цГМФ—зависимая протеинкиназа G
(PKG), подавляет активность аденилатциклазы и рецептор—зависимое
увеличение уровня Са2+ проницаемость К+ каналов и
увеличивает вход ионов К+ в клетку, способствуя гиперполяризации
плазмалеммы. Содержание цГМФ в цитоплазме регулируется разными механизмами,
одним из которых является активация фосфодиэстеразы.
В запуске фосфоинозитидной системы участвуют Gq—белки,
активирующие мембранные изоформы фосфолипазы. С (ФЛС бета) и Са2+—ионные
каналы плазмалеммы. Энзим является фосфоинозитидспецифичным и превращает
инозитолдифосфат мембраны в трифосфат. Другим продуктом гидролиза является
диацилглицерол, имеющий в боковой цепи арахидоновую кислоту. Вход Са2+
в клетку изменяет конформационную структуру рецепторов в мембранах
внутриклеточных депо катиона. При взаимодействии с инозитолтрифосфатом рецептор
приобретает функции Са2+—чувствительного Са2+—канала,
через который ион выходит из депо в цитоплазму. При увеличении концентрации
свободного Ca2+ в цитоплазме он акцептируется многими Са—связывающими
белками.
Среди Са2+—связывающих белков важен кальмодулин
(СаМ), молекула которого, акцептируя Ca2+, способна активировать специфические
киназы. Среди них наиболее полифункциональна СаСаМ—зависимая киназа II,
фосфорилирующая Na+/Н+ обменник и
играющая одну из ключевых ролей в секреторном процессе и регуляции концентрации
внутриклеточного Са2+ Воздействие ряда гормонов на тонус и
сокращение гладких и поперечнополосатых мышц через систему Gq—белков
обусловлено активацией Са—связывающих сократительных белков. Известная роль Ca2+
в регуляции рН цитозоля, транспорта и цитоскелета также подчеркивает важность
фосфоинозитидной системы, регулируемой через Gq—белки, сопряженные с
рецепторами.
Арахидонатная система вторичных посредников, связанная
с выделением из фосфолипидов мембраны и метаболизмом арахидоновой кислоты,
активируется другой группой G—белков — Gpia. Арахидоновая кислота
участвует в гиперполяризации клеточных мембран через ингибирование
деполяризующих Na+— и медленных Са2+—каналов,
а также через активацию гиперполяризующих К+—каналов. Ингибирование
GAP—белков арахидоновой и фосфатидиловой кислотами дополняет перечень их
тормозных эффектов в отношении других систем вторичных посредников.
Рецепторы III типа характерны для инсулина, факторов роста и цитокинов.
Они представляют собой рецептор и энзим одновременно, поскольку внеклеточный
домен интегральной молекулы содержит последовательность аминокислотных
остатков, связывающих определенный лиганд, а цитозольный С—конец содержит
области (домены) с киназными свойствами. Эти особенности рецептора обеспечивают
быстроту передачи сигнала о связывании гормона на внутриклеточные структуры.
Благодаря кластеризации мономерных рецепторов или же существованию стойких гомо—
или гетеродимерных молекулярных конструкций интенсивность воздействия лиганда
усиливается.
Энзиматические домены у рецепторов пролиферативных
гормонов представлены по преимуществу протеинтирозинкиназами (РТК), а у
рецепторов цитокинов могут быть и серин—треониновыми. Связывание гормона
приводит к изменению конформационной структуры и аутофосфорилированию молекулы
рецептора. Это позволяет ему затем фосфорилировать мембранные белки: энзимы,
другие типы рецепторов, ионные каналы, G—белки (соответственно по Туr или по
Ser и Thr), и изменять их активность. Кроме того, возможно
фосфорилирование цитозольных протеинкиназ с иными свойствами, а также других
энзимов и транскрипционных факторов. Совокупность этих белков представляет
систему трансдукции, обусловливающую полиэффекторные и быстрые воздействия
гормона.
В настоящее время для рецепторов факторов роста и
цитокинов также описаны системы внутриклеточных посредников.
В запуске тех или иных клеточных реакций, будь это
процессы синтеза, клеточного цикла или апоптоза, могут участвовать разные
системы внутриклеточной трансдукции сигнала. Внутриклеточные рецепторы
связывают те гормоны, которые посредством эндоцитоза, диффузии или мембранных
транспортеров могут поступать в клетку (рис. 6.34).
Гормон—рецепторный комплекс переносится в ядро и действует на синтез ДНК, изменяя скорость транскрипции и количество мРНК. Увеличение или уменьшение количества мРНК влияет на синтез белка в процессе трансляции, что приводит к изменению функциональной активности клетки.
Для оксида азота, например, гормона или трансмиттера,
характерно связывание с цитозольной гуанилатциклазой, представляющей
собой его рецептор. Цитозольные рецепторы гормонов стероидной природы,
ретиноидов и йод—содержащих тиреоидных гормонов представляют собой сложные
макромолекулы гликофосфопротеинов. Гормон—связывающий домен является
протеинкиназой, активируемой после связывания гормона и фосфорилирующей
транскрипционные факторы.
|
|
|
Рис. 6.34 Взаимодействие гормона с внутриклеточным
рецептором |
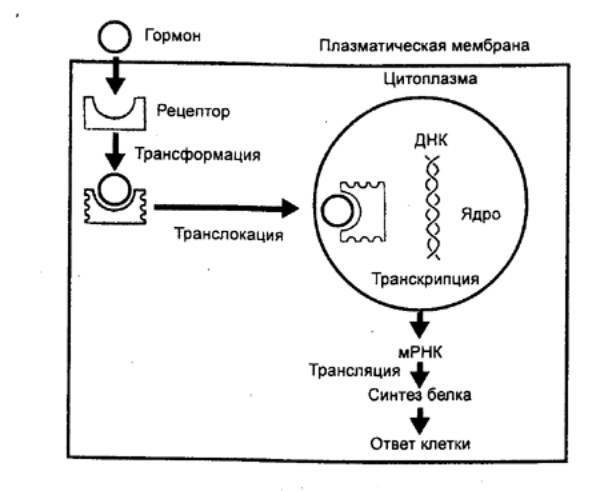
Расположенный ближе к N—концу рецептора ДНК—связывающий
домен имеет «выпетления» аминокислотных последовательностей, соединенных с
атомом цинка (так называемые «цинковые пальцы»). В цитоплазме они «закрыты» белками
теплового шока, транспортирующими лиганд—рецепторный комплекс в ядро. В
ядре цинковые пальцы обеспечивают узнавание специфического для данного
рецептора участка связывания с ДНК и взаимодействие с ним. Он представляет
собой сдвоенную (прямую или инвертированную) гормон—респонсивную нуклеотидную
последовательность в области промотора определенного гена. Чаще с ней
связывается сдвоенный лиганд—рецепторный комплекс. Следующий за этим запуск
процесса транскрипции гена может быть усилен, если образуется не гомодимер, что
характерно для стероидов, а гетеродимер (например, рецептора тироксина и цис—ретиноевой
кислоты).
Гормон—респонсивная
нуклеотидная последовательность может не только стимулировать, но и подавлять
экспрессию гена (например, рецептор глюкокортикостероидов ингибирует ген
кортиколиберина). Сходное действие описано и для мономера рецептора тироксина в
отсутствие гормона, тогда как связывание лиганда и последующая димеризация
комплекса приводит к противоположному эффекту.
Таким
образом, гормоны, как и трансмиттеры, через разные типы рецепторов систем
внутриклеточной трансдукции сигнала более или менее быстро реализуют свои
эффекты на уровне клеточной мембраны, органелл и цитоплазмы, а также ядра.
Специфика условий синтеза рецепторов и гормонов в разных тканях обусловливает
различие в них типов рецепторов к одному лиганду и, соответственно, возможность
разных эффектов.
Для поддержания гомеостаза и осуществления основных
жизненных функций в эволюции возникли две основные системы: нервная и эндокринная,
— работающие во взаимодействии между собой при осуществлении интегративной
деятельности.
Гормоны вырабатываются в
эндокринных железах, не имеющих выводных протоков, или в группах клеток с
эндокринной функцией в пределах разных органов и поступают в циркулирующие
жидкости организма. Для гормонов характерны дистантное действие и способность
вызывать специфическую активность, воздействуя на рецепторы эффекторных органов
и тканей—мишеней в низких концентрациях.
В нервной системе беспозвоночных и позвоночных
животных находятся нейросекреторные клетки, в которых вырабатываются
гормоны, регулирующие различные функции организма и стимулирующие синтез и
секрецию гормонов эндокринными железами.
У позвоночных животных регулирующее влияние ЦНС на
физиологическую активность эндокринных желез осуществляется через
нейросекреторные центры гипоталамо—гипофизарной системы.
Висцеротропные гормоны — это олигопептиды,
поступающие в основном в общую циркуляцию. Антидиуретический гормон
(вазопрессин) оказывает влияние на функцию почек, на уровень кровяного давления
и имеет ряд других эффектов, окситоцин влияет на гладкую мускулатуру матки,
секрецию молока.
Гипофизотропные гормоны (олигопептиды) поступают в
основном в ток портальной крови. Либерины (рилизинг—гормоны) оказывают
стимулирующее влияние на функцию железистых клеток гипофиза, статины
(ингибирующие факторы) тормозят их функцию. В гипоталамусе вырабатываются шесть
либеринов и три статина.
Гипофиз состоит из двух основных компонентов: нейрогипофиза и
железистой части, или аденогипофиза. В клетках аденогипофиза под действием
гипофизотропных факторов гипоталамуса вырабатываются гормоны, оказывающие
тропное действие на периферические железы.
Эффекторные гормоны (три из семи) оказывают непосредственное влияние на
органы— или ткани—мишени. К ним относятся гормон роста, влияющий на растущие
органы и ткани, пролактин, регулирующий развитие и функцию молочной железы, меланоцитостимулирующий
гормон, действующий на функцию пигментных клеток—меланофоров. Тропные
гормоны гипофиза (четыре из семи) влияют на эндокринные железы II порядка,
стимулируя их функцию. Кортикотропин регулирует синтез и секрецию
кортикостероидов корой надпочечника. Тиреотропный гормон оказывает
стимулирующее влияние на щитовидную железу, в которой происходит выработка
трийодтиронина (T3) и тироксина (Т4). Гонадотропные гормоны стимулируют
секрецию половых стероидов стероидогенной тканью гонад. Функция системы
гормональных взаимодействий осуществляется с использованием механизма обратной
связи.
Кроме гормонов, образующихся в гипофизе и в
эндокринных железах II порядка, в организме вырабатываются гормоны, синтез
которых не зависит от непосредственного регулирующего влияния гипофиза. К этим
гормонам относятся адреналин и норадреналин, синтезируемые в
мозговом веществе надпочечников, паратгормон, образующийся в
паращитовидных железах, кальцитонин, вырабатываемый в
ультимобранхиальных железах (С—клетки щитовидной железы млекопитающих), а также
ряд гормонов поджелудочной железы.
В отсутствие регуляции со стороны гипоталамуса и
гипофиза для обеспечения гомеостаза и поддержания на оптимальном уровне всех
параметров внутренней среды организма в систему регуляции включаются гормоны—антагонисты
и различные метаболиты (глюкоза, жирные кислоты, ионы и т. д.).
В желудочно—кишечном тракте вырабатываются пептиды,
выполняющие роль гормонов. В настоящее время для ряда гормонов установлена
множественная локализация синтеза, так, например, гастрин и холецистокинин,
являющиеся гормонами желудочно—кишечного тракта, обнаружены в мозгу; значение
подобных явлений требует дальнейшего анализа. В процессе эволюции гормоны, по—видимому,
возникли до образования эндокринных желез. В дальнейшем произошло развитие
сложной системы нейро—гормональных взаимодействий, в состав которой вошли
нейросекреторные элементы с вырабатываемыми ими гормонами, а также гормоны
специализированных эндокринных желез. В результате образовался единый механизм
регуляции, обеспечивающий деятельность организма.