6.5. ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА И ЕЕ ГОРМОНЫ
Поджелудочная
железа состоит из экзокринной и эндокринной частей, которые развиваются из
одного источника — энтодермы первичной кишки. В экзокринной части,
составляющей у человека 98% всей массы железы, вырабатывается пищеварительный
сок, который поступает в двенадцатиперстную кишку (см. разд. 11.2.3) и содержит
ферменты, необходимые для расщепления белков, жиров и углеводов. В эндокринной
части — панкреатических островках (Лангерганса) — синтезируются гормоны,
регулирующие метаболические процессы, в особенности углеводный обмен.
В процессе эволюции
эндокринная часть возникла раньше экзокринной, так что вначале поджелудочная
железа не имела компактного строения. У ланцетника в эпителии слизистой
оболочки кишки есть клетки, гомологичные экзокринной и эндокринной частям
поджелудочной железы. Эти клетки разобщены. У круглоротых (миноги) образуются
островки железистой ткани, отделяющиеся от эпителия кишечника. У костистых рыб
эндокринная ткань представлена островками (тельца Брокмана), обособленными от
экзокринной ткани поджелудочной железы. У других позвоночных животных эндокринная
ткань в виде мелких включений расположена среди экзокринной ткани поджелудочной
железы и составляет только 1—3% от ее массы.
Гормоны, вырабатываемые
эндокринной частью. В панкреатических
островках большинства позвоночных выявляют два основных типа железистых
клеток, вырабатывающих разные гормоны: инсулин и глюкагон.
Клетки, синтезирующие инсулин, называют (β— (или В—) клетками; клетки, вырабатывающие глюкагон,
— α— (или А—) клетками. Кроме них определен третий
тип клеток — δ—клетки, в которых синтезируется соматостатин
(рис. 6.28).
Р—клетки, синтезирующие
инсулин, обнаружены у всех позвоночных. У низших позвоночных (круглоротые,
хрящевые рыбы) α—клетки и глюкагон не
выявлены. У всех остальных позвоночных в панкреатических островках
синтезируются инсулин и глюкагон, основная функция которых состоит в регуляции
метаболических процессов; от этих гормонов в значительной степени зависит
уровень глюкозы в крови, необходимый для нормальной жизнедеятельности
организма.
Инсулин представляет собою полипептид с молекулярной массой
около 6 кДа. Он состоит из двух цепей, соединенных двумя дисульфидными
мостиками. Инсулин образуется из предшественника — проинсулина — под влиянием
протеаз. Активность проинсулина невелика (5% активности инсулина). Превращение
проинсулина в инсулин происходит в β—клетках.
Инсулин был первым полипептидным гормоном, синтезированным искусственно.
В настоящее время инсулин
(или инсулиноподобный гормон) обнаружен у многих беспозвоночных животных, что свидетельствует,
по—видимому, не только о его древности, но и важной роли в регуляции
метаболических процессов.
Глюкагон — полипептид, построенный из одной цепи с молекулярной
массой около 3,5 кДа. Кроме α—клеток
панкреатических островков глюкагон вырабатывается также в слизистой оболочке
кишечника (энтероглюкагон). Функция энтероглюкагона несколько отличается
от роли панкреатического глюкагона.
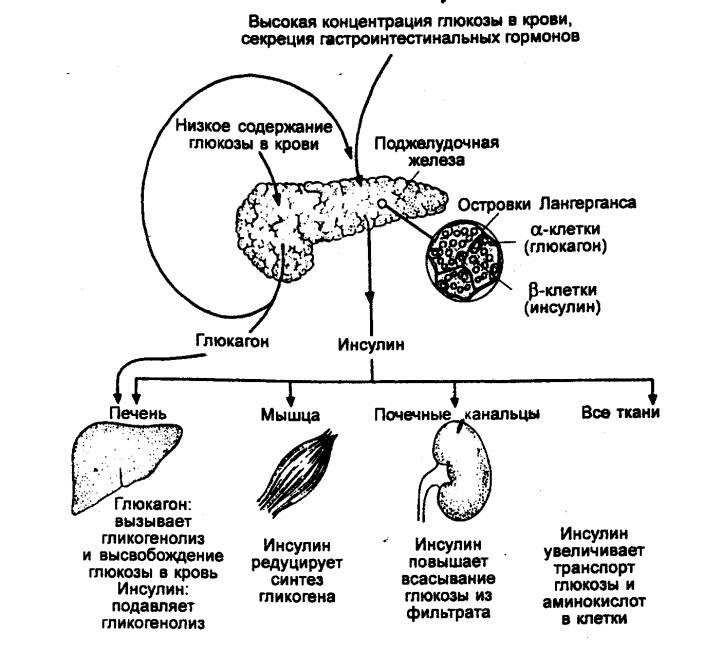
Инсулин является
анаболическим гормоном с широким спектром действия. Его роль состоит в
повышении синтеза углеводов, жиров и белков. Он стимулирует метаболизм глюкозы.
Под влиянием инсулина увеличивается проницаемость для глюкозы клеток миокарда,
скелетных мышц, что усиливает ток глюкозы внутрь клеток и ее обмен. Он
необходим для прохода аминокислот сквозь ГЭБ (см. разд. 3.27). Инсулин
стимулирует синтез гликогена в печени, снижает глюконеогенез (образование
глюкозы из аминокислот), влияет на обмен липидов, усиливая способность жировой
ткани и печени к накоплению их резервов в форме триглицеридов (рис. 6.29).
Инсулин активно взаимодействует с другими гормонами, как, например, гормоном
роста.
Действие глюкагона на
метаболические процессы осуществляется в печени и реализуется через
аденилатциклазу и цАМФ. Последний, в свою очередь, активизирует ферменты,
контролирующие скорость гликогенолиза, глюконеогенеза и липолиза. Основной
эффект гормона состоит в усилении гликогенолиза в печени, в чем глюкагон
является синергистом адреналина. По воздействию на уровень глюкозы в плазме
крови глюкагон является антагонистом инсулина.
Концентрация гормонов
поджелудочной железы в плазме крови зависит от поступления глюкозы с пищей,
скорости ее окисления и от уровня других гормонов, участвующих в регуляции
содержания глюкозы. При повышении содержания глюкозы в крови усиливается
секреция инсулина, при ее снижении
|
|
|
Рис. 6.28 Взаимоотношения между
инсулином и глюкагоном в регуляции обмена глюкозы |
|
|
|
Рис.
6.29 Действие инсулина и гормона
роста СТГ — соматотропный гормон. |

выделяется
больше глюкагона. Регуляция уровня гормона в крови осуществляется по механизму
обратной связи (рис. 6.30)
|
|
|
Рис. 6.30 Регуляция уровня гормона в крови по
механизму обратной связи |

Регуляция секреции гормонов осуществляется
симпатической и парасимпатической нервной системой. Помимо того, уровень
глюкозы, очевидно, изменяется и в самой поджелудочной железе, что используется
для регуляции секреторной активности клеток. Регуляция секреции инсулина
происходит и под влиянием ряда регуляторных пептидов, вырабатываемых в
желудочно—кишечном тракте. Присутствие в нем глюкозы вызывает выброс
энтероглюкагона. Этот гормон поступает с кровью к клеткам поджелудочной железы
и стимулирует секрецию инсулина. Инсулотропным действием обладают АКТГ,
вазопрессин и гормон роста.
Регуляция
секреции глюкагона осуществляется посредством рецепторов глюкозы в переднем гипоталамусе,
которые выявляют снижение глюкозы в крови. Возможно, в эту цепь взаимодействий включается
гормон роста гипофиза. Соматостатин, вырабатываемый δ—клетками, оказывает
ингибирующее влияние на выделение глюкагона. Энтероглюкагон также может
подавлять секрецию панкреатического глюкагона. Симпатическая стимуляция
усиливает секрецию глюкагона. Таким образом, система регуляции секреции
инсулина и глюкагона и связанного с функцией этих гормонов уровня глюкозы в
крови весьма сложна.
У млекопитающих главным источником энергии являются
углеводы, и инсулин играет весьма значительную роль в процессах регуляции их
метаболизма. У птиц в 10—20 раз более высоким является содержание глюкагона —
липолитического агента, так как основным источником энергии у этой группы
позвоночных являются липиды. При отклонениях уровня глюкозы в крови от нормы
наблюдаются явления гипо— или гипергликемии.
В
норме концентрация глюкозы в крови человека относительно постоянна и составляет
около 80—120 мг% (44,4—66,6 ммоль/л).
Патофизиология. При гипогликемии, т. е.
резком снижении уровня глюкозы, наблюдаются тахикардия, голод, возбуждение. В
результате гипогликемической комы может наступить смерть. Предотвращение этих
явлений возможно при вливаниях глюкозы и введении глюкагона. При уровне глюкозы
выше 180 мг% (99,9 ммоль/л) она выводится с мочой, что происходит при
ослабленной функции инсулина и является одним из проявлений сахарного
диабета. Это заболевание возникает в результате недостаточной выработки
инсулина β—клетками поджелудочной железы. Те же явления могут
наблюдаться в результате нарушения реакции (снижение числа рецепторов к
инсулину, снижение их аффинности (от лат. affinis — родственный, сродный) периферических тканей на
инсулин. В отсутствие гормона глюкоза медленно проникает в клетки мышц и
печени, т. к. не работают высокоэффективные инсулинзависимые транспортеры
глюкозы, запасы гликогена при этом быстро истощаются.