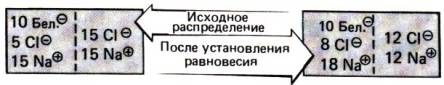
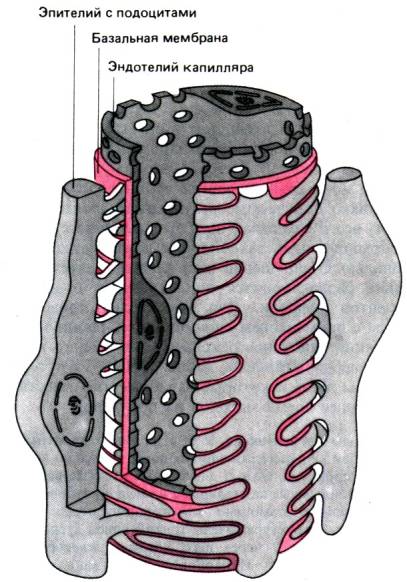
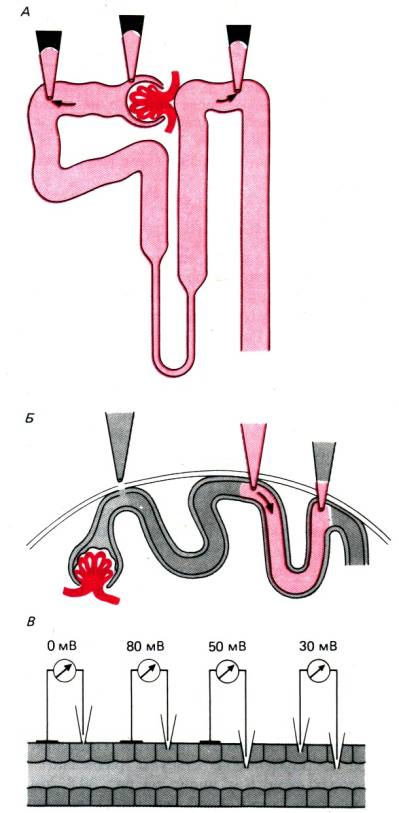
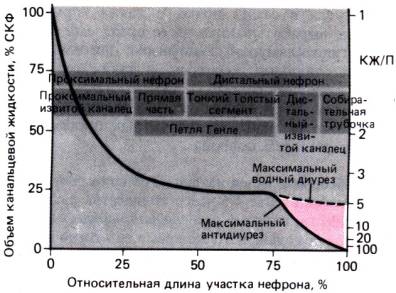
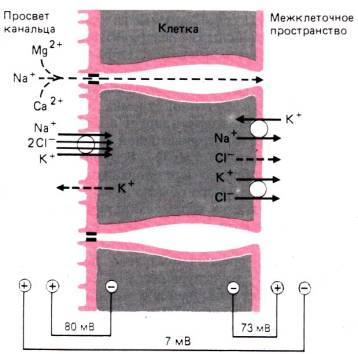
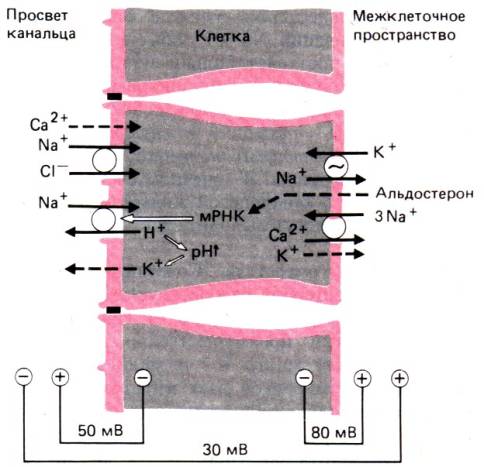
30.1. Принципы работы почек
Роль почек. Почки
поддерживают постоянство состава и объема внеклеточной жидкости,
омывающей клетки, обспечивая тем самым оптимальные условия жизнедеятельность последних.
Они выводят из организма избыток воды или растворенных в ней веществ. И
напротив, при дефиците воды и/или электролитов начинают действовать процессы,
направленные на уменьшение дальнейшей их потери без нарушения экскреции
конечных продуктов обмена.
Функциональные единицы почки. В
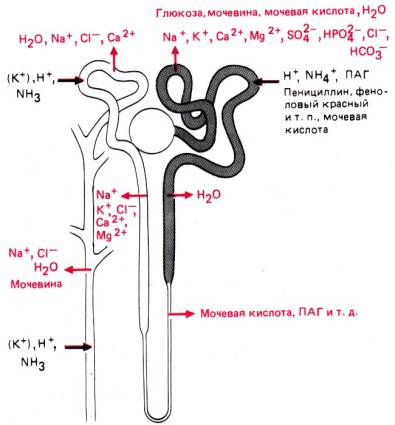
каждой почке содержится около 1,2 млн. нефронов (рис. 30.1). Отдельный
нефрон состоит из клубочка с боуменовой капсулой, проксимального извитого
канальца, петли Генле и дистального извитого канальца. Дистальные извитые канальцы
нескольких нефронов открываются в собирательную трубочку.
Каждый нефрон представляет
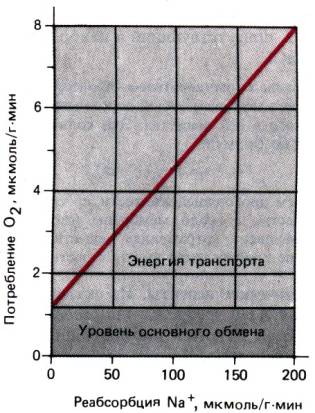
собой функциональную единицу, способную самостоятельно обеспечивать
специфические виды транспорта. Однако определенные функции почек могут
осуществляться только при совместной деятельности всех нефронов–например,
концентрирование мочи. При инактивации значительного их количества почка
утрачивает эту способность даже при нормальной работе оставшихся нефронов.
Основные механизмы функционирования почки.
В основе работы почки лежат
два главных принципа: отделение больших количеств внеклеточной жидкости от
остальной ее части путем ультрафильтрации в клубочках и транспорт
воды с растворенными в ней веществами через эпителиальные клетки канальцев. По
способности выполнять эти функции почки превзошли в процессе эволюции все
остальные органы тела. Скорость фильтрации на единицу поверхности в клубочковых
капиллярах может быть в 10 раз выше, чем в капиллярах мышц. Транспорт в
канальцах (особенно–в проксимальном извитом)–это также своего рода чудо
экономичности и эффективности.
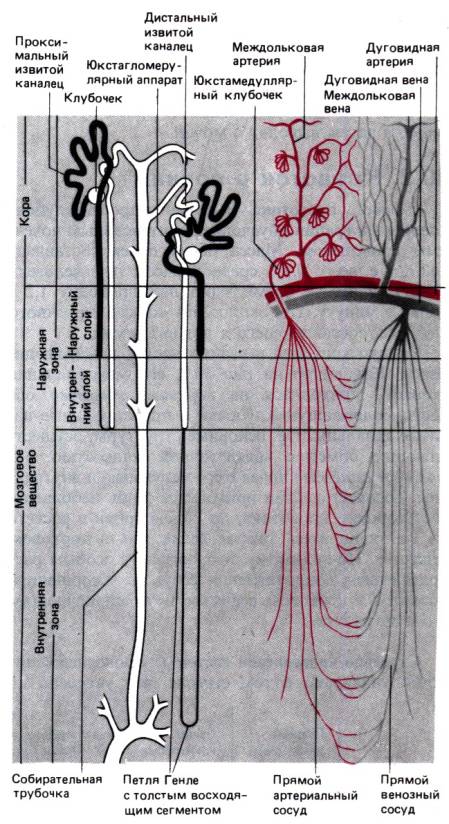
Рис. 30.1. Схема строения и
кровоснабжения нефронов. Слева изображены корковый и юкстамедуллярный
нефроны (по [19, 37] с изменениями)
Весь объем внеклеточной
жидкости, составляющий у взрослого человека не менее 17 л (см. рис. 31.1),
проходит через почки с кровотоком около 50 раз в сутки. Пятая его часть
непрерывно отфильтровывается в клубочках и поступает в канальцы. Их
клетками все вещества, которые еще могут быть использованы организмом (вода,
электролиты, витамины, аминокислоты, глюкоза и т. д.), реабсорбируются,
тогда как «бесполезные» конечные продукты обмена, а также избыток воды и
электролитов, поступивший с пищей, остаются в просвете канальцев и выводятся с
мочой. Излишек некоторых веществ (например, ионов K+ или Н+
особенно опасен для существующего в организме хрупкого равновесия. Для быстрого
и эффективного их удаления в почках помимо фильтрации существует иной механизм:
клетки канальцев извлекают эти вещества непосредственно из крови путем секреции,
а затем выводят с мочой.
30.2. Кровоток в почках
Скорость кровотока. Высокая скорость
клубочковой фильтрации обусловлена интенсивным почечным кровотоком. Масса обеих
почек составляет 300 г, т. е. всего 0,4% средней массы тела человека (70 кг). В
то же время через них проходит 1,2 л крови в минуту, т. е. около 25% общего
сердечного выброса, составляющего в покое 5 л/мин.
Однако кровоток в разных
частях почек неодинаков. Как видно на рис. 30.2, его максимальная скорость приходится на корковое
вещество–область, содержащую клубочки и проксимальные извитые канальцы, т.
е. основные структуры, осуществляющие обмен с внеклеточной жидкостью. На мозговое
вещество почек с его наружным и внутренним (сосочек) слоями приходится лишь
небольшая часть общего кровотока, но объем крови в расчете на 1 г ткани здесь
так же велик, как в корковом веществе. По–видимому, это связано с особым
распределением сопротивления сосудов, ускоряющим кровоток в корковом веществе и
ограничивающим его в мозговом.
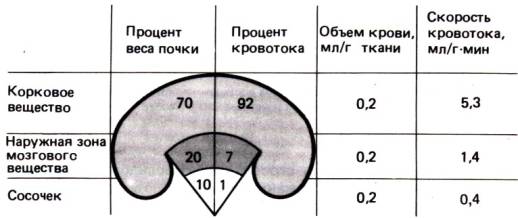
Рис. 30.2. Распределение почечного
кровотока
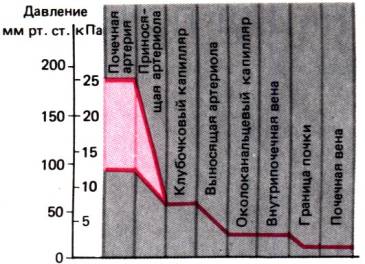
Рис. 30.3. Падение гидростатического давления на участке от
почечной артерии до почечной вены. В зоне саморегуляции (красный цвет)
за счет сопротивления приносящих артериол давление в клубочковых капиллярах
остается постоянным
Строение кровеносной системы. Кровоснабжение
почек уникально в том смысле, что устроено по принципу двух последовательных
систем сосудов с регулируемым сопротивлением и капилляров. Отходящая от
аорты почечная артерия разветвляется вблизи ворот почки на две или более
междолевые артерии. Они дают начало дуговым артериям, идущим по границе между
корковым и мозговым веществом. От этих артерий в корковое вещество отходят
междольковые артерии (рис. 30.1), разветвляющиеся на приносящие артериолы
с регулируемым сопротивлением. Те в свою очередь разделяются на плотный пучок
капилляров почечных клубочков. Из них кровь оттекает по выносящим
артериолам с регулируемым сопротивлением, которые открываются во вторую,
околоканальцевую, капиллярную сеть, окружающую извитые канальцы коркового
вещества.
Сопротивление почечному кровотоку. В условиях нормального
артериального давления его среднее падение в приносящих артериолах, т. е. на
участке от почечной артерии до клубочковых капилляров, относительно невелико
(рис. 30.3). В результате гидростатическое давление в этих капиллярах выше, чем
в любой другой капиллярной сети тела; именно оно служит движущей силой
интенсивной клубочковой фильтрации. Сопротивление выносящих артериол по
крайней мере такое же, как у приносящих.
Это обеспечивает падение
давления, достаточное для выравнивания его гидростатической составляющей в
околоканальцевом капиллярном сплетении и в проксимальном извитом канальце. В
результате жидкость из него может легко реабсорбироваться и удаляться.
Роль сопротивления приносящей
артериолы при нормальном артериальном давлении невелика, но при повышении
последнего становится очевидной. Когда артериальное давление поднимается
примерно до 200 мм рт. ст., пропорционально возрастает сопротивление приносящих
артериол и предшествующих им междольковых артерий, поэтому давление в
клубочковых капиллярах практически не меняется. Поскольку фильтрационное
давление остается постоянным, скорость фильтрации внеклеточной жидкости
также практически не меняется. Следовательно, механизмы, регулирующие ее
выведение, не могут быть перегружены.
Саморегуляция. Механизм, обеспечивающий
при колебаниях артериального давления постоянство почечного кровотока за счет
изменения сопротивления приносящих артериол, называется саморегуляцией (рис.
30.4). Такой же эффективный механизм контролирует кровоснабжение головного
мозга, где необходимо постоянство давления в постартериолярной капиллярной сети
для предотвращения отека мозга или сдавливания нервных тканей при повышении
артериального давления. Соответственно, оба этих органа по сути дела отключены
от общей системы регуляции кровообращения; в норме тонус их резистивных
сосудов не зависит от симпатического контроля и сосудистых рефлексов. Сужение почечных
сосудов может вызывать только чрезвычайно сильная симпатическая стимуляция.
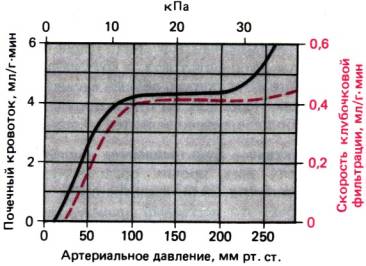
Рис. 30.4. Зависимость почечного кровотока (черная кривая)
и скорости клубочковой фильтрации (красная кривая) от артериального
давления. При давлении от 80 до 200 мм рт. ст. эти показатели почти не меняются
(саморегуляция)
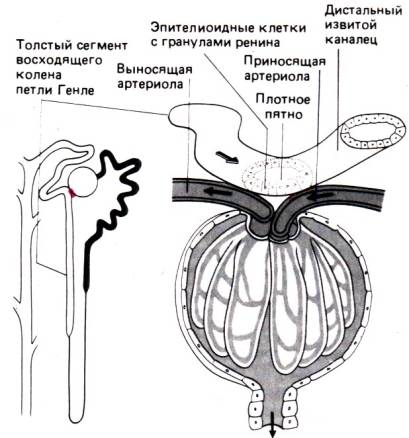
Рис. 30.5. Схема юкстагломерулярного
аппарата
Механизм саморегуляции клубочковой фильтрации и
почечного кровотока до сих пор окончательно не выяснен. Один из факторов здесь
– эффект Бейлиса (см. разд. 20.2): при повышении трансмурального
давления гладкие мышцы сосудистой стенки сокращаются. Предполагают также
существование в почках дополнительного внутреннего механизма обратной связи,
поскольку на степень сужения их артериол влияют функциональные изменения в
самих почках [45]. В основе воздействия канальца на приносящие и выносящие
артериолы лежит следующая анатомическая особенность. Конец восходящего колена
петли Генле непосредственно контактирует с сосудами в области полюса клубочка
того же нефрона (рис. 30.5). В месте этого контакта –плотного пятна (macula densa) – находится группа необычно высоких эпителиальных
клеток, а клетки в стенке артериол содержат многочисленные гранулы с ферментом
ренином. Последний представляет собой пептидазу, расщепляющую ангиотензиноген–синтезируемый
в печени белок плазмы–с образованием декапептида – ангиотензина I. Он в свою
очередь под действием другой пептидазы, называемой превращающим ферментом,
преобразуется в октапептид ангиотензин II. Последний стимулирует
секрецию корой надпочечников альдостерона (см. с, 797) и, возможно, участвует в
регуляции тонуса почечных артериол. Ангиотензин II–самое мощное из всех
известных сосудосуживающих веществ, образующихся в организме. Таким образом,
кроме механического миогенного механизма регуляции сопротивления не исключено
существование и химических агентов, снижающих скорость фильтрации.
Кровоток в мозговом веществе почек. Другая специфическая
особенность кровеносной системы почек–то, что в их мозговом веществе она
представлена только капиллярами. Последние берут начало от юкстамедуллярных
клубочков, расположенных в самом нижнем слое коркового вещества, у его
границы с мозговым. Здесь выносящие артериолы разветвляются на капилляры,
которые не образуют сплетений вокруг канальцев, а идут параллельно им, образуя
характерные пучки, направляющиеся к вершине сосочка (рис. 30.1). Эти артериальные
прямые сосуды (vasa recta) делятся на тонкие веточки,
которые впадают в восходящие венозные прямые сосуды; последние,
объединяясь в пучки, несут кровь обратно в корковое вещество. Чем дальше от
центра пучка лежит прямой сосуд, тем глубже он проникает в мозговое вещество по
направлению к вершине сосочка.
Прямые сосуды почек
достигают в длину нескольких сантиметров, тогда как в других органах тела длина
капилляров составляет около 0,5 мм. Капилляры–это участки сосудистой системы, в
которых происходит обмен веществами между кровью и внеклеточной жидкостью.
Необычно большая их длина в мозговом веществе почек, несомненно, важна для
функционирования последних. Поскольку венозные и артериальные прямые сосуды
лежат бок о бок друг к другу, а кровь по ним течет в противоположных
направлениях, на всем протяжении их контактирующих поверхностей сам кровоток
создает горизонтальный градиент концентрации всех способных к диффузии веществ.
Они соответствующим образом и диффундируют, причем те, что поступают из
канальцев в сосудистую противоточную систему (например, мочевина), все
более концентрируются по направлению к вершине петли (рис. 30.6). В случае
мочевины это важный фактор поддержания высокого осмотического давления в
мозговом веществе, обеспечивающий эффективность процессов концентрирования
мочи. Аналогичным образом накапливаются продукты, образующиеся в мозговом
веществе. Например, благодаря противоточной диффузии CО2
РCО2 здесь выше, чем в любом другом участке тела. С другой
стороны, все продукты, даже в малых количествах потребляемые в мозговом
веществе (например, О2 или глюкоза), а, значит, в нисходящем сосуде
имеющие несколько более высокую концентрацию, чем в восходящем, попадают из
первого во второй путем противоточной диффузии вблизи границы коркового и
мозгового вещества. В результате их поступление в мозговое вещество весьма
затруднено [29]. Соответственно PО2
в почечном
сосочке ниже, чем где–либо еще в организме, а довольно умеренные энергетические
потребности его ткани покрываются в основном за счет анаэробного метаболизма.
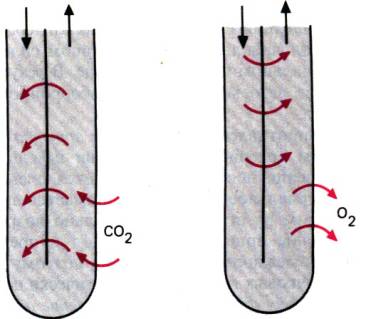
Рис. 30.6. Противоточный обмен в прямом сосуде мозгового
вещества. Образующиеся в этом слое соединения (например, СO2 ) за счет противоточной
диффузии у вершины петли Генле достигают здесь высокой концентрации. Содержание
веществ, поглощаемых в мозговом слое (например, О2), из–за
аналогичного «короткого замыкания» потоков сильно падает к вершине петли Генле
Важную роль в регуляции
кровотока в мозговом веществе, возможно, играют простагландины. В частности,
при неадекватном кровоснабжении из его интерстициальных клеток высвобождается
простагландин Е2, вызывающий расширение сосудов [15, 36, 37].
30.3. Клубочковая фильтрация
Динамика
клубочковой фильтрации
Работа фильтрующей системы. Обязательное условие
высокой скорости клубочковой фильтрации
(СКФ)–необычно интенсивный
кровоток через корковое вещество почек. Здесь непрерывно отфильтровывается
около 1/5 объема почечного плазмотока (ППТ). Иными словами,
фильтрационная фракция (отношение СКФ/ППТ) равна 0,2. За сутки образуется
170 л фильтрата, и хотя на почки приходится менее 0,5% общего объема
внеклеточной жидкости тела, через них за сутки проходит ее почти в 10 раз
больше, чем содержится во всем организме.
Эффективное фильтрационное давление. Как и в капиллярах всех
прочих органов (см. принцип Старлинга, разд. 20.6), фильтрация в клубочковых
капиллярах – это пассивный процесс, зависящий от перепада давления. Ее
«движущая сила» – разность между гидростатическим и онкотическим
(коллоидно–осмотическим) давлениями, называемая эффективным фильтрационным
давлением (ЭФД). Гидростатическому давлению в клубочковых капиллярах (Рк)
противостоит гидростатическое давление в просвете боуменовой капсулы (Рбк) и
онкотическое давление капиллярной крови (Рo)
ЭФД = Рк –
Рбк – Ро (1)
Прямые измерения методом микропункции на крысах показали, что Рк составляет 50 мм
рт.ст. (см. рис. 30.3), Рбк–12 мм рт.ст., а Ро в начальных отрезках
капилляров–20 мм рт.ст. Таким образом, эффективное фильтрационное давление
должно здесь составлять
ЭФД = 50 –
12 – 20 = 18 мм рт.ст.
На протяжении клубочковых
капилляров гидростатическое давление падает лишь незначительно (до 48 мм рт.
ст.), но поскольку из них удаляется практически безбелковый фильтрат,
концентрация белка, а следовательно, и онкотическое давление повышаются. Когда
Ро становится равным разности
между Рк и Рбк (уравнение 1), устанавливается
фильтрационное равновесие и фильтрация прекращается. Исходя из приведенных
выше цифр, такое состояние должно наступить при Ро, равном 36 мм рт. ст.,
ЭФД = 48 –
12 – 36 = 0.
Считается, что в норме
фильтрационное равновесие устанавливается еще перед концом капилляра. Значит,
при усилении плазмотока в процесс фильтрации мог бы включаться дополнительный
его участок, и благодаря увеличению фильтрующей поверхности при неизменных
фильтрационной фракции и ЭФД СКФ повышалась бы [16]. Однако действительно ли в
основе физиологических колебаний СКФ (см. ниже) лежит именно такой механизм,
пока неизвестно. Количество жидкости, фильтруемой в единицу времени за счет
градиента эффективного давления, зависит от площади фильтрации (А) и гидравлической
проводимости (С) фильтрующей мембраны:
СКФ=А·С·ЭФД,
(2)
где С – это объем воды, фильтруемый через единицу
площади мембраны в единицу времени при единичной разности давлений. Иногда его
умножают на величину поверхности (А) и полученное произведение (С • А) называют
коэффициентом фильтрации (Кф).
Физиологические колебания СКФ. Исходя из того что за сутки отфильтровывается 170 л
жидкости, CКФ в среднем составляет 120 л/мин. Это
значение часто приводят в качестве нормы. Однако следует иметь в виду, что в
течение суток происходят значительные физиологические колебания всех
параметров. Во–первых, существует четко выраженный циркадианный ритм,
максимум СКФ (дневная активность) может быть на 30% выше, а минимум (ночной
отдых)–на 30% ниже среднесуточной величины. Повышение на величину такого же
порядка наблюдается после приема пищи. Изменение количества фильтрата
может быть вызвано каждым из пяти факторов, определяющих динамику фильтрации
(уравнения 1 и 2). В патологических условиях почечная недостаточность с
дефицитом фильтрации наблюдается при изменениях давления (например, при шоке
или сужении мочеточника) или самой фильтрующей мембраны (например, при
гломерулонефрите или амилоидозе почек).
Измерение скорости фильтрации. СКФ можно измерить с помощью индикаторного
вещества по принципу Фика (см. разд. 20.13). Оно должно обладать
следующими свойствами:
1) быть фильтрующимся
неэлектролитом, т. е. не связываться с белками плазмы и не задерживаться при
прохождении через клубочковую мембрану из–за наличия электрического заряда (см.
ниже) или слишком крупного размера молекулы;
2) естественно, быть нетоксичным;
3) не расщепляться и не синтезироваться в почках;
4) не реабсорбироваться и не
секретироваться в почечных канальцах.
Любое количество такого
вещества, поступающее при фильтрации в первичную мочу, появится, не
изменившись, в моче, выделяемой из организма. Следовательно, его количество,
выделенное с мочой, равно отфильтрованному за это же время в клубочках.
Поскольку
Количество
= Объем • Концентрация, (3)
ситуация
описывается уравнением
V’м·Мв=СКФ·Пв (4)
или
СКФ= Мв/ Пв· V’м (5)
где Мв–
концентрация индикаторного вещества в моче; Пв–его концентрация в плазме, V’м–объем
мочи за единицу времени.
Одно из веществ,
удовлетворяющих указанным выше требованиям,– инулин. Это полифруктозид, состоящий
примерно из 20 остатков фруктозы, который получают из корней некоторых
фруктовых деревьев. В человеческом организме он не образуется, поэтому его
вводят внутривенно для измерения СКФ.
Определение скорости фильтрации для отдельного
нефрона. Скорость фильтрации в
отдельном клубочке можно измерить в эксперименте аналогичным способом. Обнажают
почку, вводят капилляр для микропункции
в петлю поверхностного канальца и количественно отсасывают из него
жидкость в течение определенного периода времени. Исходя из ее объема,
поступающего в каналец за единицу времени (V’к), и концентрации инулина в
фильтрате (Фин) и плазме (Пин), рассчитывают скорость фильтрации для отдельного
нефрона (СФН):
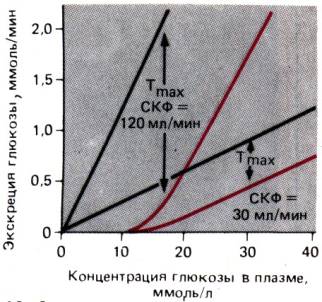
Рис. 30.18. Экскреция глюкозы при росте ее концентрации в плазме
выше пороговой величины. Разность между ее отфильтрованным (черная линия)
и выводимым с мочой в единицу времени (красная линия) количествами
отражает скорость реабсорбции. При надпороговой концентрации глюкозы эта
скорость достигает максимума (Тmax), который пропорционален СКФ
в данный момент времени
Ее концентрация,
при которой все рецепторные участки на мембране оказываются занятыми, и
является пороговой: дальнейшее повышение концентрации на реабсорбции не
отразится. В то же время котранспорт можно ускорить, увеличив натриевую
нагрузку канальцевой жидкости (см. клубочково–канальцевое равновесие). В этом
случае, несмотря на достижение пороговой концентрации, Тmax возрастает
пропорционально скорости транспорта Na+. Все это дает большое
физиологическое преимущество. Поскольку почечный порог глюкозы вдвое выше ее
нормальной концентрации в плазме, потеря ее организмом при потреблении богатой
углеводами пищи не опасна. Более того, сопряженная реабсорбция глюкозы и натрия
сводит к минимуму ее потери при повышении СКФ.
Другие углеводы. Сахароза и лактоза
подобно полифруктозиду инулину не
реабсорбируются в почках. Они свободно фильтруются и полностью выводятся с
фильтратом. В то же время дисахариды мальтоза и трегалоза
расщепляются локализованными в щеточной каемке ферментами мальтазой и
трегалазой с образованием глюкозы, которая может реабсорбироваться.
Фосфат. Фосфат реабсорбируется
почти исключительно в проксимальном канальце путем вторичного активного
котранспорта с Na+ в отношении 1:2. Это еще одна транспортная система
с типичными пороговыми признаками. В случае глюкозы почечный порог
намного превышает ее нормальную плазматическую концентрацию, и почки обычно не
играют никакой роли в регуляции последней. С другой стороны, гормоны, влияющие
на уровень глюкозы в плазме (инсулин, глюкагон, адреналин), не оказывают
непосредственного воздействия на ее транспорт в почках. С фосфатом дело обстоит
иначе. Его пороговая почечная концентрация находится в пределах нормальной
плазматической, поэтому почки играют ключевую роль в регуляции последней. Эта
их функция контролируется гормонально, в первую очередь паратгормоном
(ПГ). Он не только стимулирует мобилизацию фосфата минерального вещества
костей, но и усиливает его выведение почками, угнетая его реабсорбцию в
проксимальных канальцах с участием цАМФ. Конечный результат действия
ПГ–снижение почечного порога для фосфата [27, 33].
Сульфат также реабсорбируется в
проксимальном канальце за счет котранспорта. Однако его сродство к
транспортному белку невелико, поэтому почечный порог низок (0,8–1,2
ммоль/л). Это обусловливает низкое содержание фосфата во внеклеточной жидкости,
что важно для поддержания кислотно–щелочного равновесия. Сульфат–ион серной
кислоты (продукта расщепления белков), которая сразу после образования
нейтрализуется. Почки выделяют его в виде нейтральной соли, одновременно удаляя
избыток Н+ и возвращая организму бикарбонат, использованный в
буферных системах.
Аминокислоты. Пути аминокислот и глюкозы
в почках очень сходны. Реабсорбция аминокислот почти целиком осуществляется в проксимальном
канальце путем вторичного активного котранспорта с Na+. Транспортные
белки щеточной каемки стереоспецифичны по отношению к различным аминокислотам.
Аминокислоты с одинаковой или очень сходной пространственной конфигурацией
молекулы обладают сродством к одним и тем же рецепторным участкам, поэтому
транспортируются одним и тем же переносчиком. Пока обнаружено семь различных
систем переноса: для «кислых» аминокислот (глутаминовой, аспарагиновой); для
«основных» аминокислот (аргинина, лизина,орнитина);для «нейтральных»
аминокислот: – цистина и цистеина;– пролина, оксипролина и глицина;– глицина;–
фенилаланина, лейцина, изолейцина, триптофана и метионина; – таурина, ГАМК и (b–аланина. Врожденная или приобретенная
недостаточность одной из этих транспортных систем усиливает экскрецию с мочой
аминокислот соответствующей группы (гипераминоацидурия). При росте
плазма тической концентрации какой–либо аминокислоты насыщение специфичной для
нее транспортной системы повышает экскрецию не только ее самой, но и всех
других аминокислот, принадлежащих к той же группе [46].
Транспорт
бикарбоната и протонов
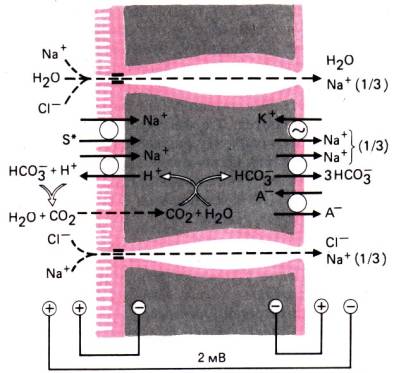
Бикарбонат. Как показано на рис. 30.13,
основная часть бикарбоната реабсорбируется из фильтрата в проксимальном
извитом канальце с помощью карбоангидразного механизма [35]. Этот
процесс зависит от секреции в просвет канальца ионов Н + за счет Na+–H+–антипорта, движущей силой
которого служит электрохимическая разность потенциалов для ионов Na+. Таким образом, поглощение
бикарбоната также сопряжено с активной реабсорбцией натрия. В результате
этого активного транспорта концентрация бикарбоната в конце проксимального
канальца снижается в норме приблизительно до 5 ммоль/л. В этом участке нефрона
существует динамическое равновесие: стенка канальца до определенной
степени проницаема для ионов бикарбоната (примерно вдвое меньше, чем для С1–),
поэтому активному их «накачиванию» в клетки противостоит обратное
«просачивание» за счет пассивной диффузии. Это равновесие выражается уравнением
Jакт=П(Cпл–Cкж) (19)
где Jакт– скорость активного всасывания;
П–проницаемость, Спл – концентрация в плазме, а Скж–равновесная концентрация в
канальцевой жидкости.
Этот механизм
«накачивания–просачивания» [55] поддерживает градиент концентрации ионов
бикарбоната на уровне около 20 ммоль/л. К концу проксимального извитого
канальца всасывается около 60% фильтрата, а концентрация в нем НСО3–
снижается до 1/5 исходной,
поэтому лишь около 8% отфильтрованного бикарбоната достигает петли Генле. Это
количество реабсорбируется главным образом в толстом сегменте ее нисходящего
колена. Однако способность к реабсорбции бикарбоната в этой части нефрона
ограничена, а в дистальном извитом канальце и собирательной трубочке она еще
меньше. Поэтому в тех случаях, когда в конце проксимального извитого канальца
концентрация НСО3– в канальцевой жидкости оказывается повышенной, эти ионы выводятся
с мочой. Результат исключительно важен: сочетание ограниченной градиентом
реабсорбции в проксимальном извитом канальце и низкой всасывающей способности
последующих участков нефрона позволяет почке регулировать кислотно–щелочное
равновесие.
Ионы водорода. В процессе обмена веществ в
организме здорового человека за сутки образуется 60–100 ммоль H+. Эти
ионы должны удаляться через почки, так
как организм способен переносить присутствие лишь следовых их количеств. При
поступлении в канальцы клубочкового фильтрата его рН равен 7,4 т. е. не
отличается от рН плазмы крови. В начальном отделе проксимального канальца он
снижается до 6,7, поддерживается на этом уровне на протяжении большей части
нефрона (рис. 30.19) и только в самом его конце падает до 5,7, а в крайне
редких случаях очень сильного ацидоза–до 4,5. Но даже при такой кислотности
концентрация ионов Н+ составляет всего 0,03 ммоль/л, т.е. в
свободном виде экскретируется только 0,05% их количества, образующегося в
процессе обмена веществ. Несмотря на столь малое содержание Н+, подкисление
канальцевой жидкости имеет исключительно важное значение. Оно необходимо не
только для реабсорбции бикарбоната, но и для регенерации бикарбонатной буферной
системы во всем организме. Более того, эта кислотность регулирует экскрецию
кислот в нейтральной форме с помощью аммиачного механизма, а также в так
называемой титруемой форме (см. ниже). Хотя почками за сутки выводится всего
0,1 ммоль протонов, способность нефронов к их секреции довольно значительна,
особенно в проксимальных извитых канальцах. Большая часть бикарбоната
реабсорбируется в буферной форме, при этом на каждую его молекулу требуется
один ион Н+. За сутки почки возвращают организму 4–4,5 моль
бикарбоната, а для нейтрализации этого количества требуется скорость секреции
протонов, которая втрое выше, чем при образовании соляной кислоты в желудке
[31, 47, 50].
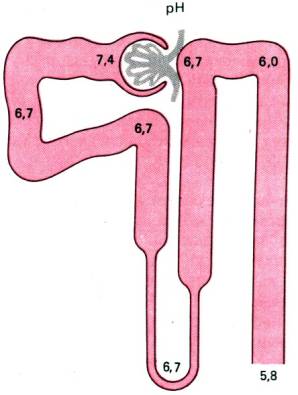
Рис. 30.19. Изменения рН канальцевой жидкости на протяжении
нефрона
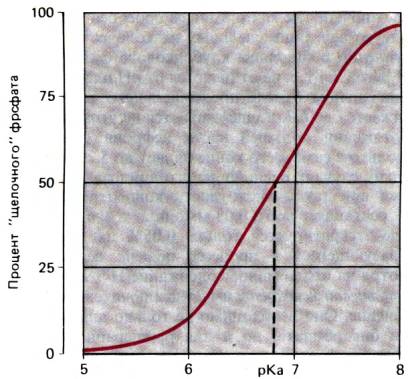
30.20. Зависимость диссоциации фосфата от рН. При кислом рН
преобладает Н2РО4–, а при щелочном–НРО42–.
При рН 6.8 (рКa) два аниона присутствуют в одинаковых количествах
Титруемые
кислоты.
«Титруемыми» называются кислоты, экскретируемые в нейтральной форме за счет
буферных соединений. Для определения их количества титруют мочу
основанием. Важнейшее буферное соединение–фосфат, который в избытке
образуется в процессе белкового и фосфолипидного обмена. Вследствие низкого почечного
порога этот избыток не реабсорбируется
из канальцевой жидкости и выводится с мочой. При рН 7,4 75% фосфата находится в
форме ионов НРО2 (вторичный фосфат), которые по мере падения рН
постепенно превращаются в НРО42– (первичный фосфат) и при рН 5,8 на 90%
переходят в эту форму (рис. 30.20).
Нейтрализуемые ионы H+ образуются при диссоциации
угольной кислоты. Связанные с фосфатом, они не просто рециркулируют, как в
случае реабсорбции бикарбоната, а выводятся с мочой. В сумме на каждый
экскретируемый Н+ заново
образуется один ион бикарбоната, и таким образом восстанавливается
содержание последнего в организме (рис. 30.21).
Аммиачный механизм. В канальцевых клетках под действием митохондриальной
глутаминазы основная аминокислота глутамин превращается сначала в глутамат–,
а затем в кетоглутарат2– [46а]. На каждом из этих этапов отщепляется
один ион аммония (NH4+). Часть этих ионов, теряя
протон, превращается в аммиак (NН3). Будучи
нейтральным, тот легко диффундирует через апикальные клеточные мембраны в
канальцевую жидкость и здесь вновь превращается в NH4+, присоединяя ионы H+,
секретируемые сюда же другим путем (Na+–H+–антипорт).
Очевидно, однако, что NH4+ может проникать через стенку
канальца и без предварительного превращения в NH3. Хотя клеточная мембрана
менее проницаема для этого иона, чем для аммиака, концентрация NH4+ в канальцевых клетках более
чем на два порядка выше. В канальцевой жидкости ионы NH4+ могут заменять Na+ при нейтрализации избытка
анионов (например, SO2–), подлежащих экскреции.
Глутарат2– в свою
очередь, взаимодействуя с двумя ионами Н+, дает СО2 и
глюкозу. Необходимые для этой реакции протоны образуются наряду с НСО3–
в реакции СО2 с Н2О,
катализируемой карбоангидразой. В конечном счете на каждый экскретируемый
ион аммония образуется один ион бикарбоната. Чем больше аммиака переходит
из клетки в канальцевую жидкость и больше образуется в ней NH4+ , тем больше секретируется H+. Однако ионы Н+
взаимодействуют и с другими присутствующими в растворе буферными соединениями, поэтому
канальцевая жидкость постепенно подкисляется. Действительно, существует почти
линейная зависимость между рН выделяемой мочи и содержанием в ней аммония (рис.
30.22). В норме аммиачный механизм выделяет за сутки в электронейтральной форме
30–50 ммоль H+ [18,44].
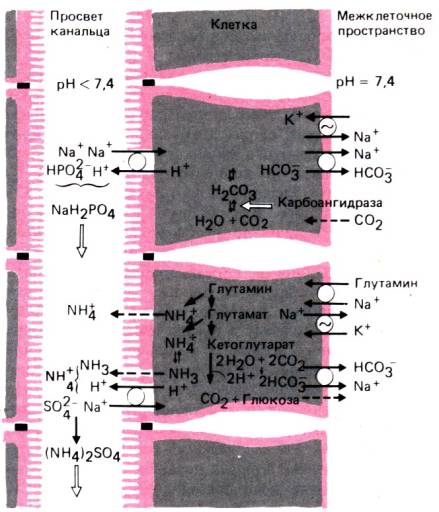
Рис. 30.21. Выделение почкой кислот. Вверху
показана экскреция титруемой кислоты, внизу аммиачный механизм
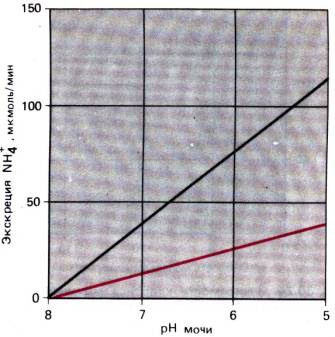
Рис. 30.22. Взаимосвязь между рН мочи и экскрецией почками
аммиака при кислотно–щелочном равновесии (красная пиния) и хроническом
метаболическом ацидозе (черная пиния) (по [5] с изменениями)
Почечная компенсация кислотно–щелочного
дисбаланса. При
необходимости (например, при диабетическом ацидозе) скорость экскреции
аммония может повыситься в 10 раз (рис. 30.22). Это обеспечивается
зависимостью от рН глутаминового обмена: при подкислении возрастает активность
глутаминазы. В результате усиливается образование NН3, удаляющего
Н+, а за счет этого восстанавливается упавшее при ацидозе
содержание бикарбоната в организме.
Не менее важную роль в
восстановлении кислотно–щелочного равновесия играет регуляция реабсорбции
НСО3–. Механизм этой регуляции определяется основными
ограничениями транспортных процессов.
Например, если
метаболический алкалоз ведет к повышению концентрации бикарбоната в плазме,
максимум, на что способны компенсаторные механизмы проксимального
канальца,–сохранение обычной разности концентраций этого иона между канальцевой
жидкостью и межклеточным пространством. Следовательно, в дистальные отделы
нефрона поступает жидкость с необычно высоким содержанием бикарбоната, а так
как его реабсорбция здесь ограничена, НСО3– выводится
с мочой (рис. 30.23, Б).
При метаболическом ацидозе концентрация бикарбоната в
плазме и канальцевой жидкости, напротив, понижена, но при сохранении
нормального градиента реабсорбируется больше НСО3– (рис. 30.23, 5). При респираторном ацидозе
повышение PCО2
снижает рН и в
клетках, так что становится больше Н+ для ионообменного
насоса, закачивающего НСО3– в клетки. В результате поддерживается более высокий
бикарбонатный градиент (рис. 30.23, Г). Таким образом, при обеих
формах ацидоза реабсорбция бикарбоната усиливается, но одновременно удаление Н+ стимулирует его ресинтез в клетках почек.
Эти механизмы продолжают действовать до тех пор, пока не восстановится
нормальное динамическое равновесие и ацидоз не будет компенсирован.
Наконец, при
респираторном алкалозе падение рсО2
приводит к повышению в клетках рН. В связи с дефицитом Н+ ослабляется ионообменная реабсорбция
бикарбоната и разность его концентраций между канальцевой жидкостью и
межклеточным пространством снижается. Избыток бикарбоната в канальцевой
жидкости выводится с мочой (рис. 30.23, Д).
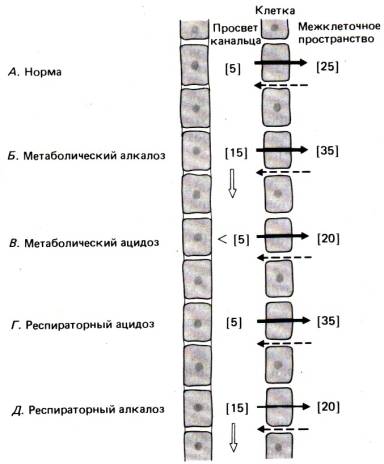
Рис. 30.23. Механизм почечной компенсации кислотно–щелочного
дисбаланса. Градиент концентрации бикарбоната, поддерживаемый за счет активного
транспорта, показан сплошными стрелками, а пассивный обратный перенос –штриховыми.
Цифры в квадратных скобках–концентрация бикарбоната (ммоль/л)
Транспорт азотистых веществ
В процессе обмена белков в
организме наряду с фосфатом, сульфатом и Н + образуются азотистые
соединения, которые также должны выводиться через почки. Два таких вещества – креатинин и аммиак уже упоминались. Кроме того, к этой категории относятся различные
соединения малой и средней молекулярной массы, среди которых наибольшее
значение имеют мочевина и мочевая кислота. В моче присутствуют
также следы белка.
Белки
и пептиды. Белки,
как уже отмечалось, в основном задерживаются клубочковым фильтром. Концентрация
их в канальцевой жидкости составляет лишь около 1% плазматической. В связи с
большим объемом фильтрата общее количество профильтровавшегося за сутки белка
доходит до нескольких граммов, но с мочой выводится не более 1% этой величины.
Реабсорбция белка происходит главным образом в проксимальном канальце. Крупные
белки поглощаются клетками путем эндоцитоза; образующиеся при этом
вакуоли сливаются с лизосомами, содержащими ферменты, которые в конечном итоге
расщепляют белки.
Пептиды (например, глутатион,
карнозин) и особенно пептидные гормоны (например, инсулин, ангиотензин,
паратгормон), молекулы которых настолько малы, что почти беспрепятственно
проходят через фильтр, так быстро расщепляются различными пептидаза.ии
щеточной каемки, что высвобождающиеся при этом аминокислоты почти полностью
реабсорбируются уже в проксимальном канальце [46].
Мочевина. У человека и большинства
позвоночных образующийся при распаде белков азот экскретируется в основном в
виде мочевины. Эта мелкая электронейтральная молекула свободно фильтруется, но
одновременно так легко диффундирует, что в проксимальном канальце около 1/3
мочевины возвращается в кровь. В дистальной части нефрона и в концевом
отделе собирательной трубочки диффузионная проницаемость для мочевины низка, но
здесь она может реабсорбироваться по механизму следования за растворителем.
Поэтому, если в дистальных отделах всасывается большое количество воды (как при
антидиурезе) и образуется моча с высокой осмотической концентрацией, вместе с
водой возвращается в кровь еще до трети отфильтрованной мочевины. В
случае водного диуреза всасывание воды в дистальном канальце прекращается, и
мочевины соответственно выводится больше. Таким образом, ее экскреция
зависит от диуреза. На рис. 30.24 это представлено в виде клиренса
мочевины. Через фильтр она проходит с такой же скоростью, как индикаторное
вещество инулин. Однако ее
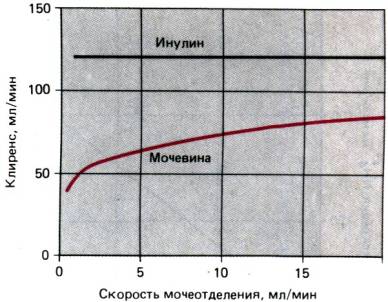
Рис. 30.24. Зависимость клиренса
мочевины от скорости образования мочи. Он сравнивается с не зависящим от
диуреза клиренсом инулина
клиренс всегда
меньше по крайней мере на 1/3. поскольку именно эта доля реабсорбируется в
проксимальном канальце независимо от диуреза. Еще 1/3 может реабсорбироваться в
дистальных отделах нефрона, когда при резко выраженном антидиурезе поглощение
здесь воды, а следовательно, и увлекаемой ею мочевины достигает максимума. С
ослаблением реабсорбции воды и увеличением диуреза экскреция мочевины растет.
Мочевина–нетоксичное,
инертное вещество, поэтому ее концентрация во внеклеточной жидкости не имеет
существенного значения и не регулируется специальным механизмом. Ее
плазматическая концентрация зависит от распада белков и СКФ (рис. 30.25).
Мочевая
кислота. В виде мочевой кислоты у
человека выводится только 5% азота. Однако это соединение имеет большое
клиническое значение, поскольку связано с возникновением подагры и
образованием почечных камней – заболеваниями, получающими в последние
десятилетия все большее распространение. Мочевая кислота–конечный продукт
обмена пуринов, поэтому ее концентрация всегда повышается, когда пища богата
компонентами клеточных ядер (т.е. включает много мяса, особенно внутренних
органов).
Мочевая кислота свободно
фильтруется. Подобно другим органическим кислотам, она также извлекается из
околоканальцевой крови клетками проксимальных каналъцев и секретируется
в канальцевую жидкость. Однако одновременно она здесь же реабсорбируется,
поэтому в конце этого отдела се примерно столько же, сколько исходно
отфильтровалось. В нисходящем колене петли Генле преобладает реабсорбция
и до вершины этой структуры доходит лишь около 10% мочевой кислоты.
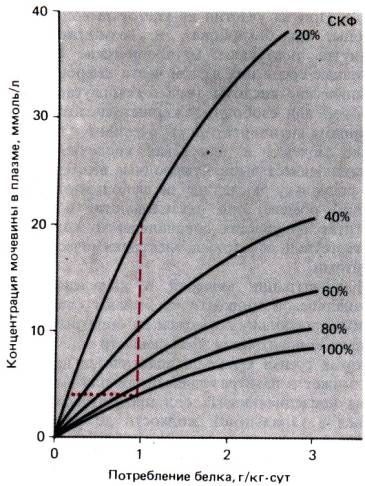
Рис. 30.25. Зависимость концентрации мочевины в плазме от СКФ и
потребления с пищей белка. При неизменном потреблении белка [штриховая
линия) концентрация мочевины возрастает, когда СКФ падает ниже нормы,
принятой за 100%. При адекватном снижении потребления белка концентрация
мочевины в плазме может оставаться «нормальной» независимо от СКФ (пунктирная
линия)
Именно это количество
выделяется с мочой, так как дистальные отделы нефрона для мочевой кислоты
практически непроницаемы (рис. 30.26). Падение концентрации мочевой кислоты еще
до того, как она пройдет через отделы нефрона, расположенные в почечном
сосочке, важно по двум причинам: ее растворимость в воде, во–первых, ограничена,
а во–вторых, зависит от рН. рКа мочевой кислоты равен 5,8, поэтому при
нормальном рН крови (7,4) она почти полностью диссоциирована до урат–аниона.
Если бы, достигнув вершины петли Генле, где самая высокая в организме
концентрация натрия, канальцевая жидкость содержала слишком много урата, то,
поскольку связанная с растворимостью величина (L)
L=[Na+]·[урат–](20)
ограничена пределом,
существовал бы постоянный риск выпадения здесь осадка.
Диссоциированная до урата, мочевая кислота в 20 раз более
растворима, чем в недиссоциированной форме. Однако канальцевая жидкость,
проходя через дистальный отдел нефрона, уменьшается в объеме и продолжает
подкисляться (рис. 30.19), поэтому концентрация мочевой кислоты возрастает, а
ее диссоциация подавляется. В собирательной трубочке, где рН равен 5,8, она на
50% не диссоциирована; при рН 4,5 эта доля возрастает до 95%. Таким образом,
реабсорбция мочевой кислоты в нисходящем колене петли Генле, т.е. до достижения
канальцевой жидкостью двух названных выше опасных участков, действует как
своего рода предохранительный клапан.
Почечные камни
Уратные камни. При нормально
функционирующих почках рН мочи редко падает ниже 5,8 (рис. 30.19). У лиц,
предрасположенных к образованию уратных камней, этот показатель часто заметно
снижен, поэтому в ней выше количество плохо растворимой мочевой кислоты (см.
выше). Кроме того, накопление и осаждение мочевой кислоты в мозговом веществе
почки и мочевыводящих путях, по–видимому, связаны с недостаточной реабсорбцией
урата в нисходящем колене петли Генле. Мочевая кислота, поступившая в прямой
сосуд, концентрировалась бы по принципу противоточной диффузии и накапливалась в вершине почечного сосочка,
если бы непрерывно не уносилась кровью. Этому транспорту способствует
связывание определенного ее количества с мембраной эритроцитов. У лиц,
предрасположенных к образованию уратных камней и развитию почечной формы
подагры (отложению мочевой кислоты в почечной ткани), связывающая способность
эритроцитов понижена.
Оксалатные камни. Щавелевая кислота–это еще
один конечный продукт обмена, который вследствие низкой растворимости в виде
имеет тенденцию осаждаться с образованием камней. Оксалат подобно
мочевой кислоте секретирустся и реабсорбируется в проксимальном канальце. В
результате его содержание в конце этого отдела
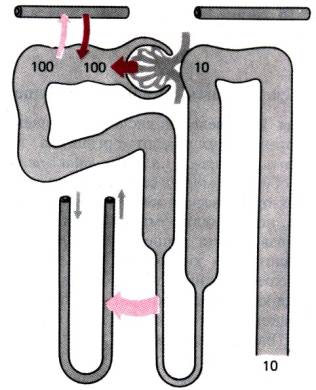
Рис. 30.26. Относительная
концентрация мочевой кислоты и
направления ее переноса в разных частях нефрона
на 25% выше количества,
отфильтрованного в клубочке. Дистальные части нефрона для оксалата почти
непроницаемы, поэтому столько же его и выводится с мочой. Концентрация его в
канальцевой жидкости, обусловленная фильтрацией и секрецией, напрямую зависит
от плазматической. В связи с этим люди, предрасположенные к образованию
оксалатных камней, должны ограничивать потребление «щавелевокислой» пищи
(например, ревеня, шпината, шоколада), а также веществ, усиливающих
метаболическое образование оксалата (например, витамина С, теофиллина).
Цистиновые камни. Цистин – наименее
растворимая из всех аминокислот. Тем не менее образование из нее камней крайне
редко обусловлено нарушением самой цистин–цистеиновой системы. Как правило, эта
патология возникает при недостаточности систем транспорта «основных»
аминокислот, когда в крови и канальцевой жидкости повышается концентрация аргинина,
лизина и орнитина. Они достаточно хорошо растворимы в воде и не выпадают в
осадок, но насыщают цистеиновую систему, к которой имеют некоторое сродство, и
вытесняют цистин из мест его реабсорбции. В результате концентрация цистина
растет, и он образует осадок.
Фосфатные
камни.
Осаждение фосфата чаще всего связано с воспалительными процессами в мозговом
веществе почек. Растворимость фосфата повышается с увеличением кислотности
среды. Если непосредственное повреждение дистальных канальцев и собирательных
трубочек нарушает местный градиент рН, канальцевая жидкость прекращает
подкисляться. К падению кислотности и осаждению фосфатов ведет также
бактериальное разложение, усиливающее выделение аммиака (действующего как
основание) в канальцевую жидкость.
Другой важный фактор –преобладание среди катионов
Са2+. Кальциевые соли обычно менее растворимы, чем, например,
натриевые, поэтому фосфат и оксалат выпадают в осадок преимущественно в виде
солей кальция.
30.13). При их участии из
клеток выводятся такие анионы, как бикарбонат, и, возможно, хлорид, а внутрь
поступают метаболические субстраты. Основные среди них в этой части
нефрона–слабые органические кислоты типа a–кетоглутаровой, фумаровой
или свободных жирных кислот. В таком анионном антипорте могут, очевидно,
участвовать также мочевая и щавелевая кислоты, а также упоминавшиеся выше
чужеродные вещества. Однако, поскольку последние не используются в клеточном
обмене, они накапливаются в цитозоле, достигая равновесия с канальцевой
жидкостью в соответствии со своими электрохимическими градиентами.
Концентрации мочевой и
щавелевой кислот в канальцевой жидкости не бывают слишком высокими, поскольку
возможен их обратный перенос. Однако концентрация здесь веществ типа ПАГ,
для которых стенка канальца практически непроницаема, может в пять раз
превышать плазматическую. Пока последняя низка, они продолжают накапливаться в
канальцевой жидкости до определенной максимальной концентрации (примерно
3,5 ммоль/л). Ее превышение невозможно, даже если плазматическая концентрация будет
расти. Как и в случае реабсорбции глюкозы, это называют «транспортным
максимумом», но такой термин опять–таки не очень удачен, поскольку под ним
часто неправильно понимают максимальное количество вещества, которое может быть
перенесено. Однако лимитирующий фактор здесь не количество, секретируемое в
единицу времени, а максимальная концентрация в канальцевой жидкости,
поэтому транспортный максимум пропорционален СКФ. Чем больше объем
жидкости, в котором растворено вещество, тем больше его накапливается при
максимальной концентрации (рис. 30.27).
Секреция экзогенных
органических веществ
Мочевая и щавелевая кислоты
отличаются от прочих органических кислот организма тем, что секретируются в
проксимальном каналъце. Однако таким же образом может секретироваться и
целый ряд слабых органических кислот, не образующихся у человека, но,
по–видимому, сходных по строению с двумя названными выше. К ним относятся
парааминогиппуровая кислота (ПАГ), рентгеноконтрастные вещества типа диодраста,
а также некоторые лекарственные препараты, например пенициллин и многие другие
антибиотики, сульфонамид, диуретики, барбитураты и др. [58].
Механизмы секреции таких соединений основаны,
очевидно, на присутствии одного или нескольких типов анионообменников на
база латеральной поверхности клеток проксимального канальца (рис.
Клиренс
ПАГ. При концентрации ПАГ ниже
максимальной до 90% ее извлекается из почечной крови путем фильтрации и,
особенно, секреции. Поскольку клиренс вещества определяется объемом плазмы, из
которого оно удаляется, у ПАГ он приблизительно пропорционален почечному
плазмотоку. Следовательно, в сочетании с гематокритом этот показатель можно
использовать для оценки скорости почечного кровотока. Раньше таков был
стандартный метод проверки функции почек, но затем его заменили более простые.
Вся ПАГ, накопившаяся в
канальцевой жидкости, выделяется с мочой, но другие органические кислоты могут
до некоторой степени диффундировать обратно. Часто в недиссоциированном виде
они имеют относительно высокую растворимость в липидах и потому легко
проходят через биологические мембраны («неионная диффузия»).
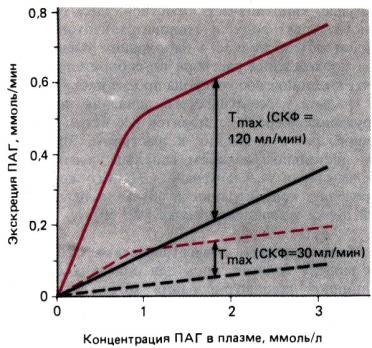
Рис. 30.27. Зависимость экскреции ПАГ от
ее концентрации в плазме и СКФ. Разность между скоростями фильтрации (черные
линии) и экскреции (красные линии) соответствует скорости секреции в
просвет канальца. При определенной концентрации ПАГ в плазме ее содержание в
канальцевой жидкости достигает максимума. При дальнейшем росте плазматической
концентрации ПАГ скорость ее переноса (Tmax) уже не повышается, но при
изменениях СКФ Tmax меняется
пропорционально
Поскольку степень диссоциации
таких слабых электролитов зависит от рН среды и их рКа, с увеличением
кислотности канальцевой жидкости усиливается их обратная диффузия. Эту ситуацию
следует учитывать в клинике. Например, в случае передозировки барбитуратов
следует как можно сильнее подщелочить канальцевую жидкость (путем введения
бикарбоната с одновременным ингибированием карбоангидразы). С другой стороны,
при лечении почечной инфекции антибиотиками нужно поддерживать максимально
высокую кислотность канальцевой жидкости, чтобы воспрепятствовать диссоциации
слабой кислоты. Это позволит добиться максимального содержания лекарства в
проникающей через мембраны форме, и таким образом повысить его концентрацию в
ткани.
30.6. Регуляция концентрации мочи
Осмолярность. Осмотическое давление
раствора зависит от количества растворенных в нем частиц; 1 осмоль
соответствует 6,06–1023 таких частиц (1 моль недиссоциирующего
вещества). Когда осмотическую концентрацию относят к объему растворителя
(осмоль/л), ее называют осмолярностью, а когда к массе растворителя
(осмоль/кг)–осмоляльностью.
Осмолярность в канальцах. При потреблении слишком
большого количества жидкости ее избыток должен выводиться. Как отмечалось выше,
в толстом сегменте восходящего колена петли Генле. NaCI
реабсорбироваться может, а вода– нет. В результате канальцевая жидкость
становится гипотоничной относительно плазмы крови: ее осмолярность (290
мосмоль/л) падает примерно в семь раз (до ≈ 40 мосмоль/л). В конце петли
Генле еще присутствует 20–25% исходного объема фильтрата (рис. 30.9). В случае
максимального водного диуреза это количество остается почти неизменным
до конца нефрона. При этом моча выделяется со скоростью 30 мл/мин и содержит
минимум осмотически активных веществ. Однако в норме основная функция почек
состоит в удалении последних, поэтому уровень водного диуреза намного ниже.
Действие АДГ.
Количество воды, реабсорбированной в дистальном отделе нефрона, а следовательно,
и уровень диуреза контролируются антидиуретическим гормоном (АДГ). Он
образуется в супраоптическом и паравентрикулярном ядрах гипоталамуса,
путем нейроаксонного транспорта поступает в заднюю долю гипофиза и там
накапливается. Высвобождение АДГ происходит постепенно в ответ на сигналы от барореиепторов
предсердий, а также периферических и центральных осморецепторов печени и
гипоталамуса (см. рис. 31.3), причем всегда в количестве, достаточном
только для обеспечения реабсорбции воды в дистальных извитых канальцах и
собирательных трубочках, поддерживающей ее нормальное содержание в организме.
Выделяется только избыточное, т.е. ненужное для поддержания гомеостаза,
количество воды [22].
Механизм концентрирования мочи
Вещества, выводимые с мочой. При смешанной диете организм
должен выделять в сутки около 7200 мосмоль отходов (конечных продуктов
обмена и т. п). Если бы требовалось, чтобы моча была изотонична плазме, на
растворение этих веществ пошло бы более 4 л воды. Однако наш водный баланс
более экономичен (и наша общественная жизнь значительно упрощена), поскольку
концентрирование мочи позволяет экскретировать ее не более 1,5 л/сутки. При
этом концентрация осмотически активных веществ в ней приблизительно втрое выше,
чем в плазме.
Для концентрирования мочи в
почках используется тот же механизм, что и для ее разведения, а именно натриевый
насос в толстом сегменте восходящего колена петли Генле и АДГ в качестве
регулирующего гормона.
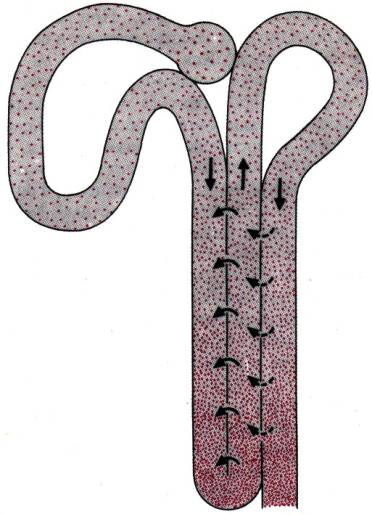
Рис.
30.28. Схема
нефрона, иллюстрирующая противоточный механизм в петле Генле. Сплошными
стрелками показан транспорт NaCI из восходящего колена в
нисходящее, а штриховыми–выход, воды из собирательной трубочки
Принцип противотока. Шпилькоообразная форма петли Генле с движением жидкости в
противоположных направлениях по лежащим рядом нисходящему и восходящему коленам
создает структурную основу концентрирования здесь мочи по принципу
противоточного умножения. Как показывает сильно упрощенная схема на рис. 30.28,
NaCI перекачивается из восходящего колена петли Генле в нисходящее, поэтому
концентрация канальцевой жидкости в последнем все более повышается. Мембрана
между двумя коленами водонепроницаема, поэтому происходит разведение раствора в
восходящем колене и его концентрирование в нисходящем. Поступающий в нисходящее
колено изотонический раствор по мере продвижения к вершине петли Генле
становится все более гипертоничным. На каждом уровне между двумя коленами
существует лишь небольшая горизонтальная разность концентраций, и в
восходящем колене натриевый насос может работать поэтапно с относительно
небольшой затратой энергии. Однако за счет противотока эти отдельные эффекты умножаются,
создавая очень большую вертикальную разность концентраций между основанием
и вершиной петли Генле. Такой градиент важен для окончательного
концентрирования мочи, и вот почему. Из восходящего колена в дистальный извитой
каналец поступает гипотонический раствор; здесь он за счет осмоса теряет воду,
и становится изотоническим, причем объем жидкости наполовину уменьшается. При
прохождении раствора по собирательной трубочке к вершине сосочка вода
продолжает удаляться по градиенту осмотического давления, поскольку окружающая
среда становится все более гипертоничной по отношению к раствору. Здесь, как и
в дистальном извитом канальце, водопроницаемость стенки регулируется
АДГ. Его количество определяет концентрацию и объем мочи, покидающей почку
в вершине сосочка [42, 60].
Модель процесса концентрирования. Столь простая модель годится
лишь для описания основных принципов концентрирования мочи, а реальные
механизмы, действующие в мозговом веществе почки, намного сложнее. Во–первых,
исключительно важна роль местного кровотока. Имеет значение также то
обстоятельство, что, хотя осмотическая концентрация максимальна в вершине
сосочка, необходимая для процесса концентрирования энергия вырабатывается не по
всей длине восходящего колена петли Генле, а только в его верхнем, толстом
сегменте. И наконец, выделяемая моча–это концентрированный раствор не NaCI, а
главным образом мочевины.
Рассмотрим схему (все еще сильно
упрощенную) на рис. 30.29 [48]. В ней допускается, что прямые сосуды в высшей
степени проницаемы для NaCI, мочевины и воды и составляют вместе с межклеточным
пространством единый жидкостный компартмент. Вслед за NaCl,
выкачиваемым из толстого сегмента восходящего колена петли Генле, в межклеточное пространство
из нисходящего колена петли Генле и соседних собирательных трубочек
устремляется вода (этот процесс регулируется АДГ). Трубочки относительно
непроницаемы для мочевины, поэтому ее концентрация в канальцевой жидкости,
достигающей внутренней зоны мозгового вещества, сильно повышается. Здесь
проницаемость для мочевины возрастает, и она диффундирует в межклеточное
пространство, что обеспечивает осмотическую потерю воды из нисходящего колена.
Последнее непроницаемо для NaCI, и концентрация этой соли постепенно повышается
к вершине сосочка. В тонком сегменте восходящего колена жидкость попадает в
противоположные по проницаемости условия: здесь проницаемость низка для воды и
высока для NaCI и мочевины. По градиенту концентрации NaCI диффундирует из
петли Генле, а мочевина поступает в нее. У толстого сегмента восходящего колена
низкая проницаемость для мочевины, как и у дистального извитого канальца и
верхнего сегмента собирательной трубочки, в которых под влиянием АДГ
реабсорбируется вода. Таким образом, концентрация мочевины в растворе
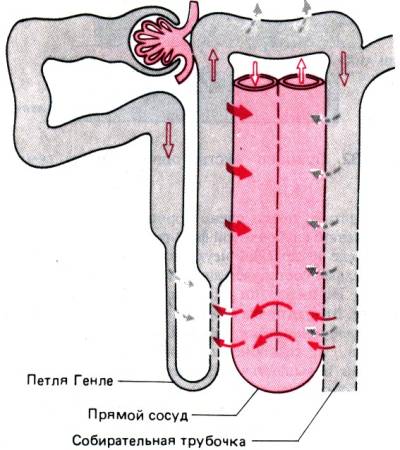
Рис. 30.29. Процесс противоточного
концентрирования мочи в мозговом веществе почек. Прямой сосуд с высокой
проницаемостью для NaCI, мочевины и воды образует единый жидкостный
компартмент с межклеточным пространством. Сюда из толстого сегмента восходящего
колена петли Генле выводится NaCI, и концентрация мочевины постепенно растет к
вершине сосочка. Структуры, проницаемые для мочевины, обозначены штриховой
линией. Белые стрелки –активны и транспорт NaCI; сплошные стрелки–
перенос мочевины; штриховые стрелки – перенос воды
непрерывно повышается, и
последняя замещает NaCI, интенсивно реабсорбируемый дистальной частью нефрона,
особенно в присутствии альдостерона. Следовательно, некоторое количество
мочевины челночно перемещается в дистальном отделе нефрона, перенося энергию
для концентрирования мочи из толстого сегмента петли во внутреннюю зону
мозгового вещества. Накоплению мочевины в мозговом веществе способствуют
прямые сосуды; в них опять–таки по принципу противоточной диффузии ее концентрация повышается к вершине петли
Генле. За счет этого эффекта, в свою очередь, поддерживается на низком уровне
градиент концентрации между кровью и жидкостью в собирательной трубочке. Итак, концентрирование
мочи, выделяемой почками, осуществляется осмотическим путем за счет транспорта
NaCl, но сам NaCl замещается мочевиной; этот конечный продукт обмена выводится растворенным
в минимальном объеме воды.
Почечная недостаточность
Важнейшее условие работы
почки–образование клубочкового фильтрата. Если его отделяется слишком мало,
состояние канальцевых механизмов реабсорбции электролитов, глюкозы или
аминокислот, а также секреции К+, Н+ или аммиака уже не
имеет значения. При недостатке фильтрата и адекватного потока жидкости в
отдельных участках канальцев все связанные с ними процессы становятся
неэффективными. Критическое сокращение объема клубочкового фильтрата
может наступать внезапно (острая почечная недостаточность) или развиваться в
процессе продолжительной болезни (хроническая почечная недостаточность).
Острая почечная
недостаточность. Причины этого функционального расстройства внепочечные. При нарушении
(нередко–всего лишь кратковременном) кровоснабжения почки, вызванном либо потерей
крови или жидкости, либо острой сердечной недостаточностью, количество
клубочкового фильтрата иногда резко падает. При этом мочеотделение также сильно
уменьшается (олигурия) или полностью прекращается (анурия). При
стабилизации кровообращения почечный кровоток обычно скоро нормализуется, но
сниженная скорость клубочковой фильтрации с олигурией или анурией могут
сохраняться. В результате вещества, выводимые в норме с мочой, задерживаются в
организме, и развивается уремия (см. ниже). Если некоторая остаточная
фильтрация еще сохраняется, часто возникает полиурия, т.е. выделение
повышенного объема мочи. В этом случае почка выводит наружу большую часть
фильтрата, почти полностью утрачивая способность к реабсорбции и секреции
электролитов, а также концентрированию или разбавлению мочи, т.е. к регуляции
водно–солевого баланса организма. Все эти нарушения через несколько дней или
недель постепенно исчезают, почти или вообще не отражаясь на морфологии почки
[12].
Острую почечную
недостаточность вызывают не только нарушения кровоснабжения, но и некоторые
яды, так называемые нефротоксины (например, аминогликозиды и тяжелые
металлы). Предполагают, что основная ее причина при этом – изменения клеточного
метаболизма [53]. В процессе аэробного обмена в клетках почек образуются агрессивные
промежуточные продукты (типа супероксидных анионов и перекисей), подлежащие
быстрой инактивации. Под действием ферментов супероксид–дисмутазы, каталазы и
глутатион–SН–пероксидазы они восстанавливаются до спиртов. Ингибирование
последнего фермента приводит к перекисному окислению липидов, а это, в свою
очередь, к разрушению мембран (рис. 30.30)–как внутренних митохондриальных, на
которых происходит синтез АТФ, так и клеточных со встроенными в них Na+–K+–АТФазой и другими
молекулярными структурами, участвующими в транспорте ионов [53]. По сравнению с
этим повреждением клеток, затрагивающим процессы образования и превращения
макроэргических соединений, необходимых для канальцевого транспорта,
уменьшение объема клубочкового фильтрата менее опасно с точки зрения
поддержания жизнедеятельности. Низкая скорость клубочковой фильтрации иногда
обусловлена механизмом обратной связи с канальцами (см. плотное пятно) [11].
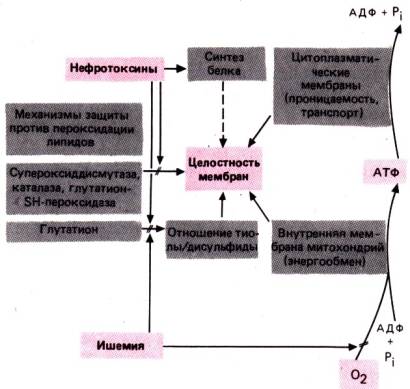
Рис. 30.30. Влияние кислородной
недостаточности или нефротоксинов на энергетический обмен и мембранные функции;
эффект связан с нарушением механизмов защиты против перекисного окисления
липидов
Хроническая
почечная недостаточность. Постепенное снижение объема клубочкового фильтрата (например, при
хроническом гломерулонефрите или закупорке кровеносных сосудов) ведет к
хронической почечной недостаточности с клинической картиной уремии.
Хотя последний термин
означает присутствие в крови избыточного количества мочевины, сама она не так
важна. Будучи конечным продуктом обмена, она практически не токсична даже в
высокой концентрации. Но повышенное содержание мочевины при уремии может
указывать на нарушение почечной экскреции других веществ, избыток которых в
жидкостях тела опасен (например, гуанидина, фенолов, различных полипсптидов и
т. п.), поскольку чреват разнообразными функциональными нарушениями работы ЦНС,
сердца, пищеварительного тракта, скелета, а также общего метаболизма и
кроветворения.
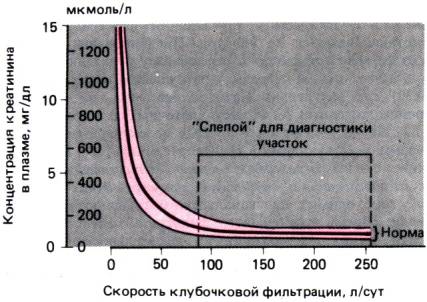
Рис. 30.31. Зависимость концентрации
креатинина плазме от скорости клубочковой фильтрации
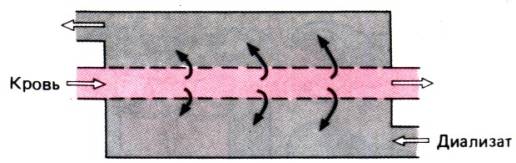
Рис. 30.32. Принцип гемодиализа.
К счастью, почки
очень хорошо приспосабливаются к ограничению клубочковой фильтрации.
Даже у здоровых людей после 40 лет СКФ неуклонно уменьшается и между 80 и 90
годами составляет лишь половину первоначальной величины. Но ни это, ни удаление
одной почки не вызывает уремии. Вторая почка или оставшиеся интактными
нефроны гипертрофируются, каждый нефрон образует больше фильтрата, а
способность канальцев к реабсорбции и секреции повышается. Прогрессирующая
почечная недостаточность развивается только после падения СКФ ниже 40% нормы.
При дальнейшем ее уменьшении внеклеточная концентрация веществ, удаляемых
только через клубочки (например, креатинина), экспоненциально повышается (рис.
30.31). Затем почки постепенно утрачивают способность выводить отходы,
участвовать в осморегуляции и поддерживать водно–солевое равновесие.
Диализ. При острой или хронической
почечной недостаточности применяют аппаратуру, позволяющую удалять из организма
конечные продукты обмена, избыток воды и солей. Старейший и самый
распространенный метод –экстракорпоральный гемодиализ: кровь из артерии
больного пропускают через систему искусственных полупроницаемых мембран и
направляют в вену (рис. 30.32). В противоположном направлении снаружи от
мембраны течет раствор, содержащий важнейшие электролиты сыворотки в тех
концентрациях, в которых они должны присутствовать в крови здорового человека.
По градиенту концентраций между кровью и диализным раствором «ненужные»
вещества переходят в диализат до тех пор, пока этот градиент не исчезнет.
30.7. Литература
Учебники
и руководства
1. Brenner
В. М., Rector F. С. (eds). The Kidney,
Vol. 1.11. W.B. Saunders Co, Philadelphia, 1976.
2. Deetjen P., Boylan J. W., Kramer K. Physiology of the Kidney
and of Water Balance, Springer, New York, Heidelberg, Berlin, 1975.
3. Greyer R., Long F., Silbemagi S. (eds.) Renal transport of
organic substrances. Springer Verlag, Berlin Heidelberg New York, 1981.
4. Massry S., Glassock R. (eds.). Textbook of Nephrology Vol. I + II, 2.
Ed., Williams + Wilkins, Baltimore, 1989.
5. Orloff J., Berliner R. W. Renal Physiology, Section 8 in
Handbook of Physiology. Am. Physiol. Soc., Washington D.C, 1973.
6. Pitts R. F. Physiology of the Kidney and of Body Fluids, Year
Book Medical Publishers, Chicago, 1972.
7. Sarre H. J., Gessler H. (eds.). Nierenkrankheiten 5. Aufl.
Georg Thieme Verlag, Stuttgart, 1986.
8. Seldin D. W., Giebisch G. (eds). The Kidney, Physiology and
Pathophysiology Vol. I, II. Raven Press. New York, 1985.
9. Siegenthaler W. (ed.). Klinische Pathophysiology 5. Aufl.
Georg Thieme Verlag, Stuttgart, 1982.
Оригинальные
статьи и обзоры
10. Baeyer van H.,
Deetjen P. Renal glucose transport. D. W. Seldin u. G. Giebisch
(eds.). The Kidney Vol. II, pp. 1663–1675, Raven Press, New York, 1985.
11. Blair–West
J.R. Renin–angiotensin system and sodium metabolism. In: K. Thurau
(ed.). Int. Rev. Physiol. Kidney and urinary tract physiology II, Vol. 11, pp.
95–143, University Park Press. Baltimore, 1976.
12. Blantz
R.C. Intrinsic Renal Failure: Acute, D. W. Seldin a. G. Giebisch
(eds.). The Kidney, Vol. II, pp. 1863–1884. Raven Press, New York, 1985.
13. Carafoli
E. The regulation of the cellular functions of Ca2 +. In: F. Bronner and J. W.
Coburn (eds.). Disorders of mineral metabolism, calcium physiology. Vol.
II, pp. 1–42. Academic Press, New York, 1982.
14. Deetjen
P., Kramer K. Die Abhangigkeit des 0;–Verbrauches der Niere von der
Na–Ruckresorption. Pfliigers Arch., Europ. J. Physiol., 273, 636–650 (1961).
15. Dunn
M.J., Hood V.L. Prostaglandins and the kidney. Am. J. Physiol., 233, F
169–184 (1977).
16. Oworkin
L. D., Brenner В. M.
Biophysical Basis of Glomerular Filtration. D. W. Seldin and G.
Giebisch (eds.). The Kidney, pp. 397–426, Raven Press, New York, 1985.
17. Giebisch
G. Renal tubular control of potassium transport. Klin. Wschr., 57,
1001–1008 (1979).
18. Goldstein
L. Ammonia production and excretion in the mammalian kidney. In: Thurau
(ed.). Int. Rev. Physio!., Vol. 11, pp. 283–316, University Press, Baltimore,
1976.
19. Gottschalk
С. W. Osmotic concentration and dilution of the
urine. Amer. J. Med., 36, 670 (1964).
20. Greger
R. Ion transport mechanism in thick ascending limb of Henle's loop of
mammalian nephron. Physiol. Rev., 65, 760 (1985).
21. Greyer
R., Schlatter E., Long F. Evidence for electroneutral sodium chloride
cotransport in the cortical thick ascending limb of Henle's loop of rabbit
kidney. Pflugers Arch., 396, 308–314 (1983).
22. Handler
J.S., OrloffJ. Antidiuretic hormone. Ann. Rev. Physiol., 43, 611–624
(1981).
23. Hierholzer
K. Sodium Reabsorption in the Distal Tubular System. In: D. W. Seldin
a. G. Giebisch (eds.). The Kidney, Vol. II, pp. 1063–1069, Raven Press,
New York, 1985.
24. Jamison
R.L., Hall D. A. Collecting duct function and sodium balance, Ann. Rev.
med., 33, 241–254 (1982).
25. Katz
A.I. Renal Na–K–ATPase: its role in tubular sodium and potassium transport.
Am. J. Physiol., 242, F 207–F 219 (1982).
26. Kerjaschki
D. Molekularpathologie des glomerularen Sialoglykoprotein Podocalyxin, dem
Haupthestandteil des "glomerularen Polyanions", in der
experimentellen und humanen glomerularen Minimalveranderung. Klin. Wschr., 63,
850–861 (1985).
27. Knox
F. G., Haramati A. Renal Regulation of Phosphate Excretion. In: D. W.
Seldin a. G. Giebisch (eds.). The Kidney, Vol. II, pp. 1381–1396,
Raven Press, New York, 1985.
28. KnoxF.G., HaasJ.A. Factors influencing renal sodium
reabsorption in volume expansion. Rev. Physiol. Biochem. Pharmacol., 92, 76–113
(1982).
29. Kramer К., Deetjen Р., Brechtelsbauer H. Gegenstromdiffusion des
Sauerstoffs im Nierenmark. Pflugers Arch. ges. Physiol., 274, 63 (1961).
30. Kramer H. J., Kruck F. Molecular basis of tubular transport
and of the action of diuretics. Klin. Wschr., 60, 1165–1263 (1982).
31. Коерреп В., Giehisch G., Malnic G. Mechanism and
Regulation of Renal Tubular Acidification. In: D. W. Seldin a. G.
Giebisch (eds.). The Kidney, Vol. II., pp. 1491–1526,ir Raven Press, New York, 1985.
32. Krilz W., Kaissling B. Structural Organization of the Mammalian
Kidney. In: D. W. Seldin a. G. Giebisch (eds.). The Kidney, Vol.
I, pp. 265–306, Raven Press, New York, 1985.
33. Long F., Greger R., Knox F., Oberleitner H. Factors
modulating the renal handling of phosphate. In: Berlyne and Thomas
(eds.). Renal Physiology, Vol. 4, pp. 1–16 (Karger, Basel, 1981).
34. Maak Т., Park С.
H., Camargo M. J. F. Renal Filtration,
Transport, and Metabolism of Proteins. In: D. W. Seldin a. G.
Giebisch (eds.). The Kidney, Vol. II, pp. 1773–1804, Raven Press, New York,
1985.
35. Maren Т.Н. Carbonic anhydrase: chemistry, physiology and inhibition. Physiol.
Rev., 47, 597–781 (1967).
36. McGiffJ.C., Wong P.Y. Prostaglandins and Renal Function.
Proc. VII Inst. Congr. NephroL, Montreal 1978, p. 83.91, S. Karger Verlag,
Basel.
37. Moffat D. В., Fourman J. The
vascular pattern of the rat kidney. J. Anat. Lond., 97, 543 (1963).
38. Murer H., Bwckhardt G. Membrane transport of anions across
epithelia of mammalian small intestine and kidney proximal tubule. Rev. Physiol.
Biochem. Pharmacol., 96, 2–51 (1983).
39. Pease D. C. Fine structure of the kidney seen by electron
microscopy. J. Histochem., 3, 295 (1955).
40. Pfaller W., Gstraunthaler G., Deetjen P. Biochemical Aspects
of Cell Injury in Acute Renal Failure. In: H. E. Eliahou (ed.). Acute
Renal Failure, pp. 25–29, John Libley, London, 1982.
41. Quamme
G.A., Dirks J.H. Magnesium: Cellular and Renal Exchanges. In: D. W.
Seldin a G. Giehisch (eds.). The Kidney, Vol. II, pp. 1269–1280,
Raven Press, New York, 1985.
42. Roy
D. R., Jamison R. L. Countercurrent System and Its Regulation. In: 0. W.
Seldin a. G. Giebisch (eds.). The Kidney, Vol. II, pp. 903–932,
Raven Press, New York, 1985.
43. Roos
A., Boron W. F. Intracellular pH. Physiol. Rev., 61, 296–443 (1981).
44. Ross
В., Lowry M. Recent developments in
renal handling of glutamine and ammonia. In: Greger, Lang, SUbernagI (eds.).
Renal transport of organic substances, pp. 78–92, Springer, Berlin Heidelberg,
New York, 1981.
45. Schnermann
J., Briggs J. Function of the Juxtaglomerular Apparatus: Local Control of
Glomerular Hemodynamics. In: D. W. Seldin a. G. Giehisch (eds.).
The Kidney, Vol. I, pp. 669–697, Raven Press, New York, 1985.
46. Silbernagi
S. Amino Acids and Oligopeptides. In: D. W. Seldin a. G. Giebisch
(eds.). The Kidney, Vol. II, pp. 1677–1702, Raven Press, New York, 1985.
47. Steinmetz
P. R. Epithelial Hydrogen Ion Transport. In: D. W. Seldin a. G. Giebisch
(eds.). The Kidney, Vol. II, pp. 1441–1458, Raven Press, New York, 1985.
48. Stephenson
J. L. Central Core Model of the Renal Counterflow System. Kidney Int., 2,
85–94 (1972).
49. Sutton
R. A. L.. Quamme G.A., Dirks J.H. Transport of calcium, magnesium and
inorganic phosphate in the kidney. In: Giebisch (ed.). Membrane
transport in biology, pp. 357–412, Springer, Berlin, Heidelberg, New York,
1979.
50. Tannen
R. L. Control of acid excretion by the kidney. Ann. Rev. Med., 31, 35–49
(1980).
51. Taylor
A., Wmdhager E.E. Cytosolic Calcium and Its Role in the Regulation of
Transepithelial Ion and Water Transport. In: D. W. Seldin a. G. Giebisch
(eds.). The Kidney, Vol. II, pp. 1297–1322, Raven Press, New York, 1985.
52. Thoenes
W., Longer K. H. Relationship between cell structure of renal tubules and
transport mechanisms. In: K. Thurau a. H. Jahrmarker (eds.).
Renal Transport and Diuretics, Berlin, Heidelberg, New York, Springer, 1969.
53. Thurau
К., Mason J., Gstramthaler G.
Experimental Acute Renal Failure. In: D. W. Seldin a. G. Giehisch
(eds.). The Kidney, Vol. II, pp. 1885 1889, Raven Press, New York, 1985.
54. Thurau
К., Wober E. Zur Lokalisation der
autoregulativen Widerstandsanderung in der Niere. Pflugers Arch. ges. Physiol.,
274, 553–566 (1963).
55. Ullrich
K.J., Fromter E., Murer H. Prinzipien des epithelialen Transportes in Niere
und Darm. Klin. Wschr., 57, 977–992 (1979).
56. Ullrich
К. J., Greger R. Approaches to the Study of
Tubule Transport Functions. In: D. W. Seldin a. G. Giebisch
(eds.). The Kidney, Vol. I, pp. 427–496, Raven Press, New York, 1985.
57. Weber
P. C., Scherer В., Siess W, Held
E.. Schnermann F. Formation and action of prostaglandins in the kidney.
Klin Wschr., 57, 1021–1030 (1979).
58. Weiner
I. M., Organic Acids and Basis and Uric Acid. In:D. W. Seldin a. G. Giebisch
(eds.). The Kidney, Vol. II, pp. 1703–1724, Raven Press, New York, 1985.
59. Windhager
E. E„ Taylor A. Regulatory role of intracellular calcium ions in epithelial
Na transport. Ann. Rev. Physiol., 45, 519–532 (1983).
60. Wrz
H., Hargitay B., Kuhn W. Lokalisation des Konzentrierungs–prozesses in der
Niere durch direkt Kryoskopie. Helv. physiol. pharmacol. Ada, 9, 196 (1951).