Глава 29
ФУНКЦИИ ЖЕЛУДОЧНО–КИШЕЧНОГО ТРАКТА
К. Эве, У. Карбах
Основное
назначение желудочно–кишечного
тракта–превращение пищи в такие молекулы, которые могут всасываться в кровь и
транспортироваться в другие органы. Начинаются эти процессы с механической
обработки пищи (измельчения, перемешивания, перемещения) и секреции
пищеварительных соков. Содержащиеся в соках ферменты расщепляют белки, жиры
и углеводы на мелкие фрагменты, способные всасываться (переваривание). Вместе
с водой, минеральными солями и витаминами конечные продукты переваривания поступают
из просвета кишечника через клетки его слизистой оболочки в кровь и лимфу
(всасывание).
Желудочно–кишечный тракт представляет собой сплошную
трубку, соединяющую ротовое отверстие с анальным, и состоит из ротовой полости,
глотки, пищевода, желудка, тонкого и толстого кишечника. В желудочно–кишечный
тракт поступают продукты секреции нескольких органов, в том числе слюнных и
поджелудочной желез и печени (рис. 29.1). Стенки пищеварительной трубки на всем
ее протяжении построены по единому плану (рис. 29.2), хотя разные ее отделы
выполняют различные функции.
Одни отделы желудочно–кишечного тракта (ротовая
полость и пищевод) служат в основном для транспортировки пищи, другие
(желудок и толстый кишечник) – для ее хранения, третьи (тонкий
кишечник)–для переваривания и всасывания. Регуляция этих функций
осуществляется 1) посредством целого ряда гормонов и биологически активных
пептидов, 2) за счет сократительной активности гладкомышечных клеток и 3)
вегетативной нервной системой. Нарушение нормальной функции пищеварительного
тракта может приводить к различным заболеваниям и клиническим проявлениям:
расстройствам пищеварения или всасывания, дискинезии (понос, запор, рвота,
недержание кала) и таким явлениям, как изжога, ощущение тяжести и переполнения,
колики и тошнота.

Рис. 29.1. Расположение органов, участвующих в переваривании и
всасывании
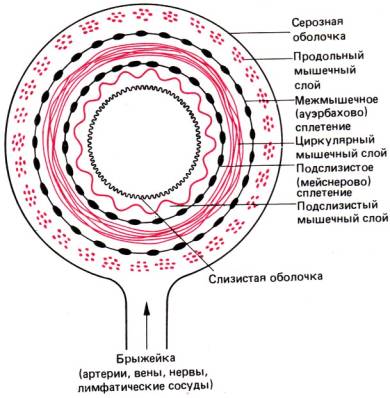
Рис. 29.2. Схематическое изображение
слоев стенки желудочно–кишечного тракта
29.1. Общие представления о функциях
желудочно–кишечного тракта
Механизмы регуляции
Внутренняя и внешняя нервные
системы . Желудочно–кишечный тракт
имеет собственную кишечную, или внутреннюю, нервную систему,
называемую также «кишечным мозгом». Она функционирует независимо от внешней
вегетативной нервной системы и регулирует моторную и секреторную активности
желудка и кишечника. Эта нервная сеть состоит из двух отделов–межмышечного
(ауэрбахова) сплетения, лежащего между слоями продольных и циркулярных
(кольцевых, круговых) мышц, и подслизистого (мейсснерова) сплетения,
расположенного между слоем циркулярных мышц и подслизистым мышечным слоем (рис.
29.2). Эфферентные волокна ауэрбахова сплетения оканчиваются на клетках гладких
мышц продольных и циркулярных слоев; они регулируют тонус мышц и ритм их
сокращений. Подслизистое сплетение регулирует в основном секреторную активность
эпителиальных клеток. Афферентные волокна обоих сплетений передают сенсорные
сигналы от механорецепторов и болевых рецепторов в центральную нервную систему.
Значительное влияние на моторную и секреторную
активности желудочно–кишечного тракта оказывает внешняя вегетативная нервная
система. Желудочно–кишечный тракт обильно снабжен парасимпатическими и
симпатическими нервными волокнами. Преганглионарные волокна блуждающего
нерва, выходящие из продолговатого мозга, иннервируют пищевод, желудок,
тонкий кишечник, проксимальный отдел толстого кишечника, печень, желчный пузырь
и поджелудочную железу, а волокна, выходящие из крестцового отдела спинного
мозга, иннервируют сигмовидную кишку, прямую кишку и область анального
отверстия. Парасимпатические нервные волокна, иннервирующие желудочно–кишечный
тракт, оканчиваются в ганглиях интрамуральных сплетений или в ганглиях,
расположенных в стенках слюнных желез и печени. Нейромедиатором в
преганглионарных нервах служит ацетилхолин, который взаимодействует с
никотиновыми рецепторами, локализованными на мембранах ганглионарных клеток.
Ацетилхолин служит медиатором и в постганглионарных нервных окончаниях, но в
этом случае он взаимодействует с мускариновыми рецепторами эффекторных клеток.
Существует множество биологически активных пептидов, также выполняющих роль
постганглионарных медиаторов; это, например, вазоактивный интестинальный
полипептид (ВИП), энкефалины, вещество Р и серотонин.
Симпатические преганглионарные нервные волокна, иннервирующие
желудочно–кишечный тракт, выходят из 5–12–го грудных и 1–3–го поясничных
сегментов спинного мозга. Они оканчиваются в чревном ганглии (пищевод,
желудок, двенадцатиперстная кишка и поджелудочная железа), верхнем
брыжеечном ганглии (тонкий кишечник и верхняя часть толстого кишечника) и нижнем
брыжеечном ганглии (нижний отдел толстого кишечника и анальное отверстие).
Нейромедиатором в преганглионарных волокнах служит ацетилхолин, а в
постганглионарных – норадреналин.
Как блуждающий, так и
симпатические нервы содержат также висцеральные афферентные волокна. Сигналы, поступающие по
этим волокнам в центральную нервную систему, участвуют в возникновении ощущений
или в запуске безусловных рефлексов.
Гормоны
и пептиды желудочно–кишечного тракта. Желудочно–кишечный тракт относится к органам,
наиболее подверженным гормональным влияниям, как по разнообразию действующих на
него гормонов, так и по диапазону эффектов. К настоящему времени в слизистой
желудочно–кишечного тракта и в поджелудочной железе обнаружено 18 видов клеток,
вырабатывающих важные для функций желудочно–кишечного тракта гормоны или
пептиды. К классическим гормонам желудочно–кишечного тракта относятся
гастрин, секретин и холецистокинин; высвобождаясь в кровь под действием
специфических стимулов, эти вещества воздействуют на определенные эффекторные
органы. В последние годы доказано также существование целого ряда биологически
активных пептидов, которые, не будучи классическими гормонами, действуют на
желудочно–кишечный тракт в основном так же, как гормоны (табл. 29.1). Некоторые
из этих пептидов действуют паракринным путем, т. е. диффундируют из
клеток, в которых они образуются, к соседним эффекторным клеткам, что не
сопровождается повышением их концентрации в сыворотке. Другие пептиды действуют
нейрокринным путем, т. е. высвобождаясь из нервных окончаний в местах их
действия. Раньше считали, что некоторые нейропептиды (энкефалины, эндорфины)
присутствуют только в мозгу, но теперь их рецепторы обнаружены и в кишечнике
[36].
Высвобождение гормонов или пептидов может
происходить при участии блуждающего нерва. Кроме того, эндокринные клетки
желудочно–кишечного тракта обладают рецепторами, которые взаимодействуют
со специфическими веществами, находящимися в просвете кишечника. Под действием
этих веществ из базальных частей клеток высвобождаются гранулы с гормонами,
поступающими затем в капилляры. Регуляция образования гормонов в
желудочно–кишечном тракте отличается от таковой в других эндокринных системах
тем, что секреция гормонов зависит не столько от концентрации
Таблица 29.1. Гормоны и биологически активные пептиды
желудочно–кишечного тракта
|
Гормоны
|
Основные функции
|
|
Гастрин
|
Стимулирует желудочную секрецию, вызывает
трофические эффекты
|
|
Секретин
|
Стимулирует панкретическую секрецию (бикарбонат)
|
|
Холецистокинин
|
Стимулирует панкреатическую
секрецию (ферменты), сокращения желчного пузыря
|
|
Биологически активные
пептиды («кандидаты» в гормоны)
|
Соматостатин
|
Угнетает секрецию (желудок,
поджелудочная железа)
|
|
Панкреатический полипептид
|
Угнетает секрецию
(панкреатического сока, желчи)
|
|
Урогастрон
|
Угнетает секрецию (желудок)
|
|
Энтероглюкагон
|
Угнетает секрецию (желудок,
поджелудочная железа). Стимулирует желчеотделение
|
|
Нейротензин
|
Угнетает секрецию и
опорожнение желудка, вызывает сужение сосудов
|
|
ГИП (глюкозозависимый
инсулинотропный пептид)
|
Вызывает высвобождение
инсулина
|
|
Нейропептиды
|
|
ВИП(вазоактивный
интестинальный полипептид)
|
Угнетает желудочную секрецию, стимулирует
панкреатическую секрецию (бикарбоната) и независимое от желчных кислот
желчеотделение. Расслабляет гладкие мышцы
|
|
Вещество Р
|
Стимулирует слюнные железы
и сокращение гладких мышц
|
|
Энкефалины, эндорфины
|
Угнетают сокращения гладких
мышц
|
|
|
|
гормонов или пептидов в
крови, сколько от прямого взаимодействия компонентов пищи с эндокринными
клетками пищеварительного тракта.
Гормоны и пептиды желудочно–кишечного тракта можно
разделить на две группы по их аминокислотным последовательностям. К первой
группе относятся гастрин и холецистокинин, имеющие пять
одинаковых концевых аминокислотных остатков. Оба соединения действуют на один и
тот же рецептор на поверхности клетки и с одинаковым результатом, хотя
эффективность их действия может быть разной в зависимости от специфичности
рецептора. Так, например, гастрин влияет на париетальные клетки сильнее, чем
холецистокинин, а последний оказывает более сильное действие на сокращения
желчного пузыря по сравнению с гастрином. Наиболее характерным представителем второй
группы гормонов и пептидов является секретин. В эту же группу входят
вазоактивный интестинальный полипептид, глюкагон и глюкозозависимый
инсулинотропный пептид – все родственные секретину по наличию в
полипептидной цепи одной и той же
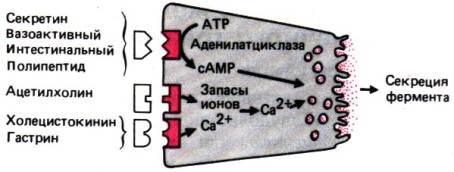
Рис. 29.3. Стимуляция секреции
ферментов различными гормонами и медиаторами и внутриклеточные механизмы их
действия (сопряжение стимуляции и секреции)
аминокислотной
последовательности. В одних случаях гормоны этих двух групп действуют как
антагонисты по отношению к эффектам друг друга, однако их эффекты могут быть и
синергическими в определенном органе. Примером синергизма может служить
действие этих гормонов на ацинозные клетки поджелудочной железы (рис. 29.3).
Секрецию ферментов поджелудочной железой стимулируют как гормоны первой группы
(гастрин и холецистокинин), так, хотя и в меньшей степени, некоторые гормоны
второй группы (секретин и ВИП). В результате взаимодействия всех этих «первых
посредников» с их рецепторами повышается внутриклеточная концентрация
кальция (гастрин, холецистокинин) или увеличивается образование цАМФ (секретин
и ВИП). В свою очередь последние, действуя в качестве «вторых посредников»,
стимулируют секрецию ферментов клетками поджелудочной железы.
Моторика желудочно–кишечного тракта
Виды перистальтики. Переваривание и всасывание в
желудочно–кишечном тракте в большой степени зависят от изменений конфигурации
его стенок, связанных с сокращением и расслаблением их мускулатуры. Основные
виды перистальтики показаны на рис. 29.4. Продвижение пищевого комка от
орального отверстия к анальному обусловлено пропульсивной перистальтикой
волнообразно распространяющимися по кишке сокращениями циркулярных мышц,
которым, как правило, предшествует волна расслабления. Перемешиванию
пищевого комка с пищеварительными соками способствуют непропульсивная
перистальтика, распространяющаяся лишь на небольшое расстояние, и
ритмическая сегментация. Сегментация означает одновременное сокращение циркулярных
мышц то в одних, то в других близко расположенных участках кишки,
чередующихся с участками расслабления. Поскольку при этом частота сокращений
уменьшается в направлении от верхних отделов кишечника к нижним, содержимое
кишечника благодаря непропульсивной перистальтике медленно перемещается в
сторону анального отверстия.
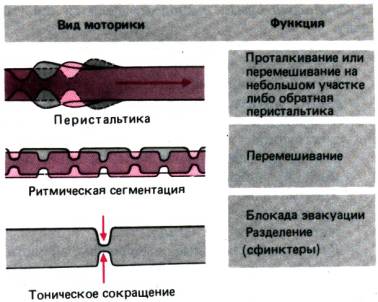
Рис. 29.4. Виды перистальтики
желудочно–кишечного тракта и их функциональное значение
Различные в функциональном
отношении отделы пищеварительного тракта отделены друг от друга за счет
тонического сокращения и периодического расслабления специализированных
участков–сфинктеров. Например, между пищеводом и желудком находится нижний
сфинктер пищевода, а между подвздошной кишкой и слепой – илеоцекальная
(баугиниева) заслонка. Благодаря сокращению этих сфинктеров пища продвигается
только в одном направлении.
Регуляция перистальтики. Потенциал покоя на мембранах гладкомышечных клеток
желудочно–кишечного тракта характеризуется спонтанной ритмической
деполяризацией, носящей название медленных волн. Эта деполяризация
не вызывает механического ответа мышц, но если на нее накладываются
кратковременные потенциалы действия, то в результате входа в клетки ионов Са2+
развивается мышечное сокращение. Сила сокращений зависит от числа потенциалов
действия. В связи с этим каждое сокращение мышц коррелирует с возникновением
медленной волны. Основной ритм медленных волн варьирует в разных отделах
желудочно–кишечного тракта и составляет в желудке, в двенадцатиперстной кишке и
подвздошной кишке соответственно 3, 12 и 8 сокращений в минуту.
«Голодный» мноэлектрический двигательный комплекс. Когда в желудке и тонком
кишечнике иссякают остатки пищи, в пищеварительном тракте происходит характерное
явление – ритмические сокращения, получившие название «голодных». После
относительно продолжительного периода покоя (фаза 1; длительность около
1 ч) и эпизодических –сокращений (фаза 2, продолжительность около 30
мин) резко повышается электрическая и двигательная активность (фаза 3;
продолжительность около 15 мин). Эти фазы показаны на рис. 29.5. Во время фазы
3 в антральном отделе желудка или в двенадцатиперстной кишке возникают
многочисленные потенциалы действия и сильные сокращения, частота которых
составляет 10–12/мин. Фронт «голодных» сокращений распространяется вниз
по тонкому кишечнику, и когда он достигает подвздошной кишки, в верхнем отделе
желудочно–кишечного тракта возникает новая волна сокращений. Скорость продвижения
фронта «голодных» сокращений составляет 6–8 см/мин в верхнем отделе кишечника и
2 см/мин в нижнем отделе, а весь цикл занимает около 1,5 ч. Фронт активности
проталкивает вперед остатки пищи, скопления бактерий и даже чужеродные
предметы. Этот перемещающийся комплекс двигательной активности получил образное
название «домоправителя» желудочно–кишечного тракта. У больных с патологически
высоким количеством бактерий в тонкой кишке эта двигательная активность
нарушена.
Миоэлектрический двигательный комплекс возникает в гладкомышечных
клетках и в интрамуральном нервном сплетении кишечника. Возникает он
автономно, но может подвергаться модифицирующему воздействию со стороны
вегетативной нервной системы или гормонов. Особую роль в этом процессе играет
гормон мотилин, основным источником которого служит слизистая верхних
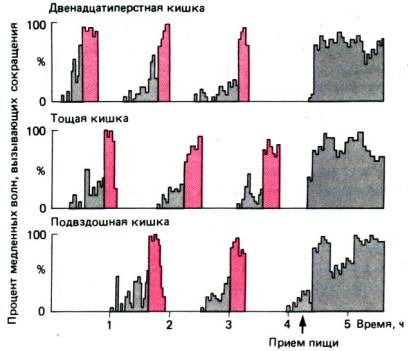
Рис. 29.5. «Голодный» Миоэлектрический
двигательный комплекс. Процент медленных волн, за которыми следуют «голодные»
сокращения мышц. В фазе 1 двигательная активность отсутствует. В фазе 2 (серые
столбики) 50% медленных волн уже вызывают сокращения, а в фазе 3 (красные
столбики) сократительная активность достигает максимума. Фронт двигательной
активности перемещается от двенадцатиперстной кишки (верхний рисунок) и
подвздошной (нижний рисунок) за 1,5ч, и затем весь цикл начинается
сначала. Прием пищи (отмечен стрелкой) прерывает этот «голодный»
комплекс
отделов тонкого кишечника. Об этом свидетельствует
повышение концентрации мотилина в крови в фазе 3, а также тот факт, что весь
комплекс может быть запущен преждевременно путем внутривенного введения
мотилина.
Основные механизмы секреции
Пищеварительные соки интенсивно синтезируются в
секреторных клетках различных органов–слюнных желез, ротовой полости,
желудочных и кишечных желез и экзокринного отдела поджелудочной железы.
Выделяемый этими клетками первичный секрет представляет собой раствор,
содержащий минеральные соли, ферменты и другие белки. При прохождении
первичного секрета через выводные протоки его состав, особенно электролитный,
может подвергаться значительным изменениям.
Функциональная анатомия. Пищеварительные
ферменты–это белки, образующиеся в специализированных железистых клетках.
Их синтез начинается в шероховатом эндоплазматическом ретикулуме
базальной части клетки. По цистернам или канальцам эндоплазматического
ретикулума новообразованный белок достигает апикальной части клетки и попадает
в цистерны аппарата Гольджи, где он концентрируется в зимогеновых
гранулах диаметром около 3 мкм. Под действием секреторных стимулов гранулы
высвобождаются из клетки путем экзоцитоза. При этом мембрана гранулы сливается
с поверхностной мембраной апикальной части клетки, и в месте слияния образуется
отверстие, через которое содержимое гранулы выделяется из клетки в проток.
Регуляция секреции. Активность пищеварительных желез может
стимулироваться блуждающим нервом и соответственно угнетаться атропином.
Кроме того, секрецию желез активируют или подавляют специфические гормоны.
В качестве вторых посредников в этом процессе участвуют ионы Са2+ и
цАМФ (см. рис. 29.3).
Иммунная система,
связанная с желудочно–кишечным трактом
Вместе с усвояемыми
компонентами пищи и неперевариваемыми волокнистыми компонентами в
пищеварительный тракт попадает множество антигенов – бактерий, вирусов и
пищевых аллергенов. Для защиты от них в желудочно–кишечном тракте
имеется иммунокомпетентная лимфоидная ткань. Эта ткань составляет около
25% всей его слизистой оболочки, и общий объем ее равен почти половине объема
селезенки. В анатомическом и функциональном отношении лимфоидная ткань,
связанная с
желудочно–кишечным трактом, делится на три отдела.
1. Пейеровы бляшки– скопления лимфоидных
фолликулов, в которых собираются попадающие в кишечник антигены и
вырабатываются антитела к ним.
2. Лимфоциты и плазматические клетки, присутствующие
в собственной пластинке и образующие иммуноглобулины, главным образом IgA;
поскольку эти иммуноглобулины содержат две полипептидные цепи большего размера,
чем IgA в сыворотке, их называют секреторными IgA (sIgA).
3. Внутриэпителиальные
лимфоциты, представленные в основном Т–лимфоцитами.
Частью иммунной системы
желудочно–кишечного тракта можно считать также мезентериалъные лимфатические
узелки и ретикулоэндотелиальную систему печени, которые служат
продолжением дренажной системы лимфы и портальной крови.
Три указанные системы
находятся в непосредственном контакте с антигенами, попадающими в кишечник.
Внутриэпителиальные лимфоциты отделены от просвета кишечника только
плотными контактами между эпителиальными клетками. Между пейеровыми бляшками и
просветом кишечника находятся специальные клетки, называемые М–клетками; они
способствуют транспорту антигенов в лимфатические фолликулы. Наконец,
лимфатические клетки собственной пластинки расположены рядом с капиллярами и
лимфатическими сосудами (см. рис. 29.32). В норме этот иммунологический барьер
обеспечивает достаточную защиту, но при инфекции слизистой, вызванной особо
патогенными бактериями, или под действием других вредных факторов он может быть
разрушен.
Газы желудочно–кишечного
тракта
Газы желудочно–кишечного
тракта изучены недостаточно, а между тем их скопление в кишечнике (метеоризм)
имеет важное клиническое значение.
Объем и состав газов. Объем газов в
организме может быть измерен с помощью плетизмографии–метода, основанного на
вдувании в кишечник аргона. Количество газа, выделяющегося из кишечника, можно
определить с помощью введенного в кишечник катетера. В норме в
желудочно–кишечном тракте содержится менее 200 мл газов. Ежедневно через прямую
кишку выделяется около 600 мл газов (от 200 до 2000 мл), и выходят они в виде
приблизительно 15 порций по 40 мл каждая, хотя существуют значительные
индивидуальные различия. На диете, богатой целлюлозой, количество газов может
сильно увеличиваться за счет расщепления целлюлозы бактериями в толстом кишечнике.
При употреблении в пищу бобов количество выделяемого газа может увеличиваться в
десять раз. Однако чувство распирания кишечника не всегда коррелирует со
скоплением в нем газов. Когда субъекту с синдромом раздражения толстой кишки
вдувают инертный газ или раздувают введенный в кишечник резиновый баллон, он
испытывает чувство распирания значительно раньше, чем здоровый человек. У таких
больных понижен порог внутрикишечного давления.
Газовая смесь кишечника на 99% состоит из N2, O2 , СO2 , Н2 и СН4.
Относительные пропорции этих газов сильно колеблются в зависимости от
индивидуальных особенностей человека и происхождения газовой смеси. Неприятный
запах выделяемых из кишечника газов связан с присутствием следовых количеств
таких ароматических соединений, как индол, скатол, меркаптан и сероводород.
Происхождение
газов. Существуют три основных
источника газов в кишечнике: заглатываемый воздух, газы, образующиеся в
просвете кишечника, и газы, диффундирующие из крови.
Газовый «желудочный пузырь» образуется в результате
заглатывания воздуха. При каждом акте глотания воздух попадает в желудок;
его количество варьирует у разных людей, но в среднем составляет около 2–3 мл.
Большая часть этого воздуха выходит из желудка обратно при отрыжке. В
просвете самого кишечника образуются СO2 , Н2 и СН4.
Диоксид углерода образуется в результате реакции между ионами НСО3-,
секретируемыми поджелудочной железой, кишечником и печенью, и ионами Н+,
выделяемыми с желудочным соком, а также отщепляемыми от жирных кислот. Таким
путем образуются большие количества СO2 (РCО2 в двенадцатиперстной кишке составляет
200–500 мм рт. ст.), но значительная его часть реабсорбируется в тонком
кишечнике. СO2 в составе
газа, скапливающегося в толстом кишечнике, образуется в результате
бактериального разложения углеводов. При бактериальном разложении
невсасывающихся углеводов в толстом кишечнике образуется также Н2.
Животные, рожденные в стерильных условиях, и новорожденные дети не выделяют Н2.
По той же причине в тонком кишечнике, содержащем очень мало бактерий (см. рис.
29.40), Н2 практически не образуется.
Часть H2, образующегося в толстом
кишечнике, всасывается в кровь, поступает с нею в легкие и удаляется с
выдыхаемым воздухом. Это явление используют в широко, применяемом рутинном
тесте на скорость эвакуации пищи из тонкого кишечника. Обследуемый принимает
плохо всасывающийся углевод, после чего с помощью газовой хроматографии
анализируют выдыхаемый воздух и фиксируют время, прошедшее между приемом углевода
и повышением содержания Н2 в выдыхаемом воздухе. Эта проба
используется также для выявления нарушения всасывания углеводов.
Метан (СН4), так же как Н2,
образуется в результате разложения углеводов в толстом кишечнике. Этот процесс,
очевидно, зависит от состава кишечной флоры. Приблизительно у 1/3 взрослых
людей образуются довольно большие количества СН4, поэтому плотность
фекальных масс составляет у них менее 1,0 и они всплывают на поверхность воды.
Еще одним источником газов в
просвете кишечника служит их диффузия из плазмы. Направление диффузии
определяется относительными парциальными давлениями газа в плазме и в просвете
кишечника. В плазме давление N2 (PN2, = 600 мм рт. ст.), O2 (PО2 – 50 мм рт.ст.) и СO2 (натощак, когда низко содержание НСОз– и Н2)
выше, чем в просвете кишечника, поэтому газы диффундируют в кишечник. Диффузия N2 в кишечнике составляет 1–2
мл/мин, а объемы диффузии O2 и СO2 очень малы
вследствие низких значений парциального давления этих газов в плазме.
Н2 и СН4 образуют
с О2 взрывчатую смесь. Имеются сообщения о взрывах, приводящих
иногда к смертельному исходу, которые возникали во время колоноскопического
удаления полипов с помощью высокочастотной диатермии после вдувания воздуха,
если кишечник был неполностью очищен или применялся маннитол, который
расщепляется бактериями.
Основные механизмы
транспорта в кишечнике
Основная функция кишечника –всасывание
воды, минеральных солей и продуктов переваривания пищи, и в этих
процессах тонкий кишечник и толстый кишечник выполняют разные функции. Основная
функция тонкого кишечника– транспорт энергетического материала, воды,
минеральных солей, желчных кислот и витаминов. Толстый кишечник служит
резервуаром для каловых масс, а также играет важную роль в регуляции конечного
всасывания жидкости из кишечника. Несмотря на функциональные и анатомические
различия между тонким кишечником и толстым, механизмы транспорта в их эпителии
в принципе одинаковы.
Терминология. Содержимое кишечника (химус),
перемещается в двух направлениях–от орального отверстия к анальному
(благодаря сократительной активности стенок кишечника) и от слизистой
поверхности к серозной, т.е. из просвета кишечника в эпителий и далее в
субэпителиальные капилляры и лимфатические сосуды. Здесь мы рассмотрим
последние из названных транспортных процессов, используя понятие скорость
транспорта, или поток.
В принципе вещество может переноситься через
эпителий кишечника в любом направлении, т.е. из просвета кишечника к серозной
поверхности и наоборот. Когда преобладает однонаправленный транспорт от
слизистой оболочки к серозной, происходит процесс всасывания, а когда
перемещение происходит от серозной поверхности к слизистой, т.е. в просвет
кишечника, имеет место секреция (рис. 29.6). Конечный эффект всегда является
результирующей двух разнонаправленных потоков.
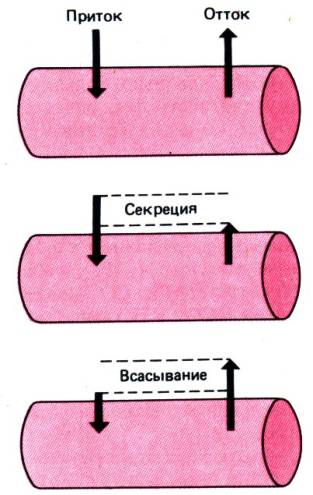
Рис. 29.6. Перемещение воды и
минеральных солей в кишечнике. Суммарный перенос является равнодействующей двух
противоположных потоков. Если преобладает поступление в просвет кишечника, речь
идет о секреции. Когда выход из просвета кишечника превышает поступление в
него, имеет место всасывание. При одинаковой интенсивности обоих процессов
суммарный результат равен нулю
Методы изучения транспорта в кишечнике. Для изучения транспорта
веществ в кишечнике используют методы как in vivo, так
и in vitro. Всасывание в кишечнике
человека можно исследовать с помощью балансовых методов, метода перфузии и так
называемых тестов на толерантность. При использовании балансовых методов
определяют разницу между количеством вещества, принятого через рот, и
количеством того вещества, которое выделяется с калом и мочой; эта разница
показывает усвояемость данного вещества в кишечнике.
Перфузию кишечника производят через многоствольные катетеры,
вводимые в кишечник через рот. Перфузионный раствор вводят в кишечник через
проксимальное отверстие катетера и отбирают через дистальное отверстие. По
разности в содержании вещества в исходном растворе и в аспирате оценивают
эффективность его всасывания.
Клиническая проба на толерантность заключается
во введении исследуемого вещества через рот и последующем измерении содержания
этого вещества или его метаболита в моче (d–ксилозная проба) или в
выдыхаемом воздухе (проба на Н2 в выдыхаемом воздухе). Эти
методы не дают, однако, никакого представления о механизмах транспорта;
для их исследования необходимо определять
однонаправленные потоки.
Механизм транспорта вещества, имеющего электрический
заряд, может быть точно охарактеризован только при исключении электрохимических
градиентов, а осуществить это можно в условиях in vitro с
помощью камеры Азинга (рис. 29.7). При таких исследованиях для
регистрации потоков вещества в том и другом направлениях используют
радиоактивные изотопы, а электрохимические градиенты устраняют, замыкая электрическую
цепь через эпителий путем приложения извне электрического напряжения.
Для анализа отдельных стадий переноса веществ
необходимо «открыть» эпителий, считающийся до сих пор «черным ящиком». Для
этого существуют два пути: измерение электрических параметров внутри
клеток с помощью записывающих электродов, встроенных в интактную ткань, и
изучение процесса транспорта в изолированных клеточных органеллах путем
выделения транспортных везикул из слизистой и серозной поверхностей
клеточной мембраны.
Функциональная анатомия
энтероцитов. Два
соседних энтероцита (клетки кишечного эпителия) и заключенное между ними
межклеточное пространство составляют функциональную единицу (рис. 29.8).
Со стороны, противоположной просвету кишечника, энтероциты и межклеточное
пространство ограничены базальной мембраной. Измерение силы тока
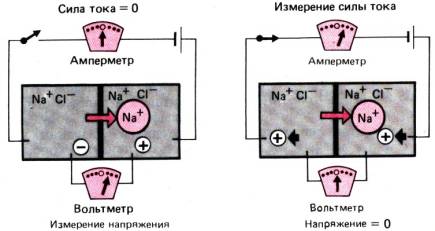
Рис. 29.7. Измерение in vitro
однонаправленных потоков с помощью «камеры Азинга». Фрагмент эпителия
закрепляют между двумя камерами, заполненными газированным раствором
электролита, и с помощью двух электродов, расположенных на слизистой и серозной
поверхностях, регистрируют разность спонтанных трансмембранных потенциалов
слизистой. Разность потенциалов уравновешивают с помощью приложенного извне
электрического тока; таким образом, цепь через эпителий замыкается (по [13])
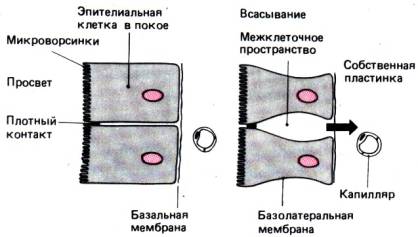
Рис. 29.8. Эпителиальные клетки в покое
и во время всасывания. Соседние клетки образуют вместе с плотным контактом и
межклеточным пространством функциональную единицу. Форма эпителиальных клеток и
межклеточного пространства зависит от функционального состояния эпителия
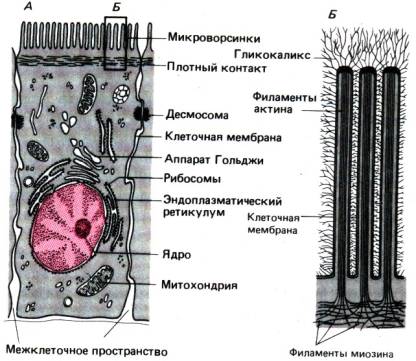
Рис. 29.9. Строение энтероцита (А)
и микроструктура щеточной каемки (Б)
Последняя не играет большой роли
в направленных процессах транспорта, поскольку ее структура такова, что через
нее могут свободно проходить даже крупные молекулы. На поверхности
энтероцитов, обращенной в просвет кишечника, имеются микроворсинки. С этой
стороны соседние энтероциты соединены плотными контактами, получившими
свое название из–за того, что под микроскопом они имеют вид плотных структур.
Первоначально считали, что плотные контакты являются барьером для диффузии, но
теперь известно, что они по крайней мере частично проницаемы для воды и
растворенных малых молекул, а также играют особо важную роль в процессах
транспорта в верхних отделах желудочно–кишечного тракта [24].
Форма энтероцитов и межклеточного пространства
подвергается заметным изменениям в зависимости от их функционального состояния
(рис. 29.8). Когда желудок пуст, энтероциты плотно примыкают друг к
другу и разделены таким узким межклеточным пространством, что его с трудом
удается различить под микроскопом. Во время процесса всасывания объем
энтероцитов уменьшается вследствие увеличения межклеточного пространства за
счет поступления в него жидкости и повышения гидростатического давления. Это межклеточное,
гидростатическое давление служит движущей силой для транспорта воды и
минеральных солей из межклеточного пространства в субэпителиальные капилляры и
лимфатические сосуды.
Клетки слизистой содержат дифференцированные системы
органелл (рис. 29.9). Эндоплазматический ретикулум играет важную роль в
синтезе белка; в этом компартменте, в частности, синтезируются белковые компоненты
хиломикронов, образующихся при всасывании жиров, и многие носители,
облегчающие транспорт веществ через клетку. В аппарате Гольджи хранятся
и подвергаются химической модификации всосавшиеся, а также новосинтезированные
в клетке вещества. В энтероцитах присутствуют также лизосомоподобные
структуры, в которых происходит гидролитическое расщепление всосавшихся и
синтезированных в самой клетке веществ. Обилие митохондрий в энтероцитах
указывает на высокую интенсивность окислительного обмена, необходимого для
выполнения энтероцитами их транспортной функции.
Характерная особенность энтероцитов, имеющая
исключительно важное значение для процессов всасывания,–это наличие
микроворсинок–пальцевидных выростов на поверхности клеток, обращенной в
просвет кишечника (рис. 29.9). Микроворсинки образуют щеточную каемку
энтероцитов шириной около 1–2мкм. Каждая микроворсинка представляет собой
цилиндрический вырост цитоплазмы, внутри которого имеется сократительная
структура, состоящая из филаментов актина и обеспечивающая изменение
формы микроворсинок. На апикальной стороне энтероцита волокна разветвляются,
образуя сложную сеть (терминальное сплетение). Как правило, плотность
микроворсинок уменьшается в кишечнике в направлении от орального отверстия к
анальному и колеблется от 650 до 3500 на клетку. В тонком кишечнике
микроворсинки покрыты дополнительным волокнистым слоем–гликокаликсом,
компоненты которого синтезируются и секретируются на поверхность энтероцитами.
В гликокаликсе присутствуют пищеварительные ферменты, адсорбированные
или образованные в самих энтероцитах; в основании слоя гликокаликса эти
ферменты встроены в мембрану микроворсинки. Сам гликокаликс покрыт тонким слоем
воды, который не удаляется даже при сильных перистальтических сокращениях; его
называют «неперемешивающимся водным слоем». Этот слой препятствует
прохождению липофильных веществ и облегчает перенос растворимых гидрофильных
частиц.
Внутри– и межклеточный пути транспорта. Вся поверхность энтероцита
покрыта трехслойной мембраной (см. гл. 1). Химический состав мембраны,
обращенной в просвет кишечника, обеспечивает транспорт через нее
жирорастворимых веществ путем простой диффузии. В то же время транспорт липидов
через водную среду цитоплазмы и базолатеральную мембрану требует затрат энергии
и осуществляется при участии особого механизма.
Перенос полярных и
несущих электрический заряд веществ через липидную мембрану происходит
чрезвычайно медленно. Чтобы дать приемлемое объяснение имеющимся
экспериментальным данным, необходимо постулировать существование в мембране пор,
или каналов, хотя до сих пор их не удалось обнаружить с помощью
светового микроскопа. Для пассивного транспорта заряженных веществ важное
значение имеет наличие на поверхности энтероцитов фиксированного отрицательного
заряда.
Наряду с переносом веществ
через клетки имеет место их транспорт через плотные контакты и межклеточное
пространство. Поскольку проницаемость эпителия определяется в основном плотными
контактами, его физические и электрические свойства во многом зависят от этих межклеточных
структур.
При электронной микроскопии с
использованием техники замораживания–скалывания плотные контакты имеют вид
нитей, сплошь заполняющих промежутки между клетками. Число нитей в плотных
контактах уменьшается в направлении от орального отверстия к анальному. Однако
плотность этих контактов зависит, по–видимому, не только от числа нитей, как
думали раньше, но и от их состава [17].
Свойства
эпителия.
Транспорт веществ через эпителий кишечника почти на 90% (в разных отделах
кишечника по–разному) осуществляется не через энтероциты, а межклеточным
путем. Проникновение веществ через эпителий межклеточным путем за счет
осмотического градиента называется пассивной проницаемостью. Плотные
контакты в тонком кишечнике полностью проницаемы для молекул диаметром до 0,8
нм и мало или совсем непроницаемы для более крупных молекул. Таким образом, для
более крупных молекул эпителий служит полупроницаемой мембраной и ведет
себя как сито, разделяющее частицы разных размеров.
Если вещество, неспособное проникать через эпителий,
поместить в растворе гипертонической концентрации на одну поверхность эпителия,
то осмотический градиент (тс) вызовет приток воды со стороны другой его
поверхности. Величина осмотического давления пропорциональна разности
концентраций маркера (вещества сравнения, неспособного проникать через
мембрану):
p=D[S]·R·T,
(1)
где D [S] – разность концентраций, R газовая
постоянная, Т–абсолютная температура.
Исходя из осмотического градиента, создаваемого в
результате присутствия вещества на одной стороне эпителия, можно рассчитать
проницаемость последнего для этого вещества. Отношение тока воды, вызванного
осмотическим давлением вещества, не проникающего через мембрану, к току воды,
вызванному исследуемым веществом, называют коэффициентом отражения. Этот
коэффициент служит мерой пассивной проницаемости эпителия для незаряженных
водорастворимых веществ (рис. 29.10). Он равен нулю в случае вещества, для
которого эпителий полностью проницаем, и единице в случае маркера, для которого
он непроницаем. Если коэффициент отражения меньше 1, но больше 0, вещество
частично диффундирует через мембрану. В этом случае размер молекул вещества
сравним с диаметром пор плотных контактов.
Диаметр пор, или каналов, плотных контактов
уменьшается вдоль кишечного тракта в направлении от проксимальных отделов к
дистальным. Эпителий тощей кишки человека проницаем для молекул диаметром
0,75–0,8 нм, подвздошной кишки –0,3–0,5 нм, а толстой–только 0,22–0,25 нм.
Проницаемость эпителия для воды в разных участках кишечника поэтому тоже
различна. При введении гипертонического раствора в просвет двенадцатиперстной
кишки приток воды в него в единицу времени будет в 3 раза больше, чем в толстой
кишке (см. также рис. 29.34). Однако пассивная проницаемость заряженных
частиц зависит не только от диаметра каналов, но и от заряда частиц.
Следует также отметить, что диаметр ионов зависит от их конфигурации, гидратной
оболочки и атомного веса.
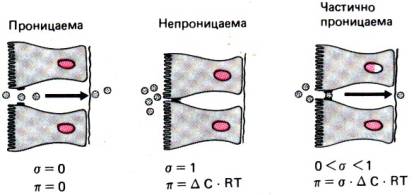
Рис. 29.10. Схема пассивного транспорта через эпителий. Перенос воды
через эпителий происходит под действием осмотического давления, создаваемого
растворенным в ней веществом; проницаемость мембраны для исследуемого вещества
может быть рассчитана путем сравнения вызываемого этим веществом тока воды с
током воды, вызываемым маркером–веществом, для которого мембрана полностью
непроницаема. Рассчитанная таким образом проницаемость называется коэффициентом
отражения σ (по [13])
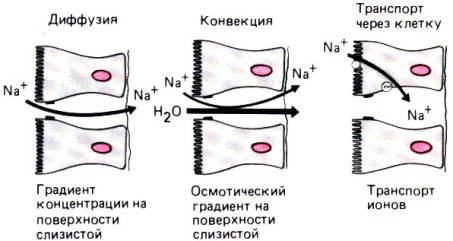
Рис. 29.11. Механизмы транспорта, участвующие в процессах
всасывания
Поверхностный заряд эпителиальной клетки.
Благодаря химическому составу плазматической
мембраны поверхность эпителиальной клетки обладает фиксированным
отрицательным зарядом. С отрицательно заряженными группами поверхности
взаимодействуют катионы (мобильные катионы), и таким образом
поддерживается электронейтральность. На поверхности мембраны, обращенной в
просвет кишечника, к таким катионам относится H+, поэтому рН на границе
между апикальной поверхностью энтероцита и просветом кишечника ниже, чем в
самом просвете (эффективная зона рН). Благодаря наличию этой зоны
ионизированные основания (например, лекарственные препараты) накапливаются на
поверхности эпителия, обращенной в просвет кишечника, что облегчает их
всасывание.
Избирательная проницаемость плотных контактов для
ионов. Экспериментальные данные
убедительно свидетельствуют о том, что катионы проходят через межклеточное
пространство в эпителии легче, чем анионы, а это означает, что плотные
контакты также несут фиксированный отрицательный заряд [41] (рис. 29.11).
Таким образом, эпителий ведет себя подобно катиончувствительной мембране, и
проницаемость межклеточного пространства зависит не только от размера молекул,
но и от их заряда. Так, например, при разной концентрации растворов NaCl на
двух сторонах эпителия ионы Na+ будут диффундировать в
сторону меньшей его концентрации активнее, чем ионы С1–. Благодаря
такой избирательной проницаемости плотных контактов, обеспечивающей
более легкую диффузию ионов Nа+, создается
трансэпителиальная разность потенциалов (диффузионный потенциал, рис.
29.11). Разность потенциалов, обусловленная диффузией иона в направлении более
низкой концентрации, описывается уравнением Нернста. Однако потенциал,
измеряемый в действительности, бывает меньше рассчитанного по уравнению, а это
означает, что плотные контакты должны быть хотя бы частично проницаемы
для ионов С1–. Относительную величину пассивной проницаемости двух
ионов можно рассчитать по уравнению Голдмана. Согласно этому расчету, пассивная
проницаемость для ионов натрия в ободочной кишке в 7 раз, а в эпителии желчного
пузыря в 3 раза больше, чем для ионов хлора.
Другой вид транспорта, зависящий от избирательной
проницаемости плотных контактов,–это следование за растворителем, или
объемный транспорт (поток). Данный вид транспорта обусловлен тем, что при
перемещении в межклеточном пространстве (конвекция) вода увлекает за
собой растворенные в ней вещества. При этом вследствие избирательной
проницаемости плотных контактов катионов переносится больше, чем анионов, т.е.
потенциал потока также способствует образованию трансэпителиального
потенциала. Диффузионный потенциал и потенциал потока возникают за счет
пассивных процессов. Величина, а также положительное или отрицательное значение
этих потенциалов зависят от осмотического и гидростатического градиентов и
трансэпителиальных градиентов концентрации электролитов. Поскольку направления
этих градиентов варьируют, величина потенциалов также колеблется.
Трансэпителиальная разность потенциалов. Путем регистрации
потенциалов на обеих сторонах эпителия тонкого и толстого кишечника можно
показать, что трансэпителиальная разность потенциалов существует даже в том
случае, если диффузионные и осмотические силы исключены. Возникновение этого
потенциала связано с процессом активного энергозависимого транспорта,
поэтому он называется транспортным потенциалом (рис. 29.11). В данном
случае эпителий работает подобно батарее, заряд которой поддерживается в
основном за счет активного транспорта натрия в направлении от слизистой
поверхности к серозной. При таком направлении транспорта натрия серозная
поверхность эпителия приобретает положительный заряд по отношению к слизистой
поверхности. Трансэпителиальная разность потенциалов увеличивается в
орально–анальном направлении, и в двенадцатиперстной кишке равна 3 мВ, а в
сигмовидной и прямой –40 мВ.
Транспорт натрия, необходимый для создания
трансэпителиальной разности потенциалов, можно изучать, замыкая электрическую
цепь через эпителий путем приложения электрического напряжения (рис. 29.7). По
закону Ома электрическое сопротивление эпителия может быть рассчитано, исходя
из спонтанной трансэпителиальной разности потенциалов и силы тока в цепи.
Электрическое сопротивление эпителия кишечника повышается в
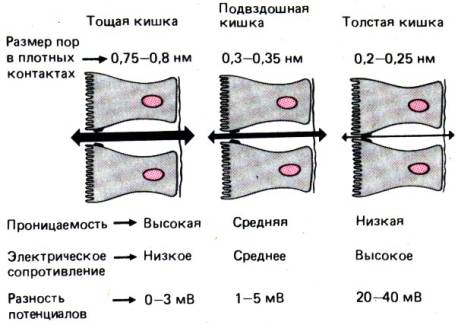
Рис. 29.12. Пассивный перенос через эпителий
зависит от размера пор плотных контактов, который уменьшается в направлении от
проксимальных отделов кишечника к дистальным, в связи с чем в том же
направлении увеличиваются трансэпителиальная разность потенциалов и
электрическое сопротивление
направлении от
проксимального отдела к дистальному. В тощей кишке оно составляет 25, а в
толстой 100–200 Ом/см2. Сама эпителиальная клетка обладает очень
большим сопротивлением, и проведенные недавно опыты показали, что сопротивление
эпителия целиком определяется межклеточным шунтом, а именно плотностью
плотных контактов [41]. Иными словами, электрическое сопротивление эпителия
обратно пропорционально размеру пор плотных контактов (рис. 29.12). В тонком
кишечнике пассивная проницаемость эпителия относительно высока, а его
электрическое сопротивление соответственно мало, благодаря чему он обладает
значительной проницаемостью. Эпителий толстого кишечника, напротив, является
относительно плотным, поэтому его электрическое сопротивление довольно
значительно. Таким образом, поглощение натрия в большой степени зависит от
сопротивления плотных контактов.
Пути и механизмы транспорта в кишечнике
Существуют два пути
транспорта воды и растворенных в ней веществ через эпителий кишечника:
транспорт через эпителиальные клетки и через межклеточное
пространство. В первом случае это активный процесс, а во втором–пассивный.
Пассивный перенос через
межклеточное пространство. Перенос веществ через межклеточное пространство всегда является
пассивным процессом и основан на диффузии или конвекции. Диффузия–это
перемещение веществ через эпителий в направлении электрохимического или
химического градиента. Необходимым же условием для переноса веществ путем конвекции
является трансэпителиальный ток воды под действием осмотических или гидростатических
сил. Как следует из названия, пассивный перенос осуществляется без затраты
клеткой энергии; он всегда пропорционален осмотическому или концентрационному
градиенту и может быть описан уравнением Фика. Всасывание воды и минеральных
солей через внеклеточное пространство в количественном отношении имеет особенно
большое значение в проксимальных отделах кишечника, т. е. в «протекающем»
эпителии (см. рис. 29.12).
Активный перенос через
эпителий.
Чтобы вещество могло всосаться через эпителиальную клетку, оно должно
проникнуть в нее через плазматическую мембрану, обращенную в просвет кишечника,
пройти через цитоплазму и выйти через базолатеральную мембрану в межклеточное
пространство (рис. 29.13 и 29.15). Совершенно очевидно, что этот ряд барьеров
значительно тормозит пассивный перенос через энтероцит. В частности, для
гидрофильных и отрицательно заряженных веществ липидная мембрана служит главным
препятствием. Важная функция эпителиальной клетки состоит в активном
транспорте веществ из просвета кишечника в субэпителиальные капилляры и
лимфатические сосуды. Характерная особенность активного переноса заключается в
том, что при этом вещество перемещается «в гору», т.е. против градиента
концентрации даже при низкой концентрации этого вещества в просвете
кишечника. Активный перенос осуществляется при участии переносчика и
требует затраты энергии. Процесс характеризуется кинетикой насыщения, т. е. его
скорость не может быть выше определенного уровня (рис. 29.14). Кроме того,
механизмы активного переноса обладают высокой степенью субстратной
специфичности и могут угнетаться веществами, имеющими химическое сходство с
субстратами, а также рядом других веществ. Нередко система активного транспорта
работает только при
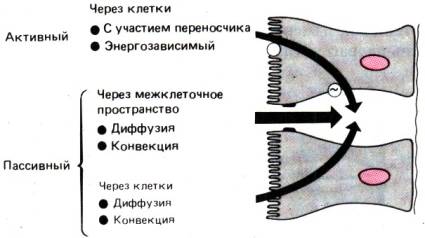
Рис. 29.13. Пути и механизмы транспорта
воды и растворенных в ней веществ через эпителий. В транспорте через клетки
участвуют и пассивный, и активный процессы, а в транспорте через межклеточное
пространство–только пассивный (по [13])
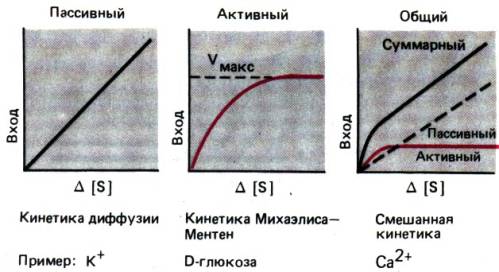
Рис. 29.14. Кинетика процессов пассивной
диффузии и активного транспорта с участием переносчика
мобилизации переносчика
другим веществом (сопряженный транспорт). Следует сказать, что процесс, идущий
с участием переносчика, не всегда отвечает всем критериям активного транспорта.
Так, например, фруктоза переносится механизмом с участием переносчика,
действующим по принципу насыщения, но без затраты энергии, а лишь за счет
разности концентраций субстрата. Такой вид транспорта называется облегченной
диффузией.
Для многих веществ характерен
транспорт смешанного типа, т.е. при участии как активного, так и пассивного
процессов. При низкой концентрации вещества преобладает активный транспорт, а
при более высоких концентрациях, когда процесс активного транспорта достигает
насыщения, преобладает процесс пассивного переноса (рис. 29.14).
Еще один механизм поглощения
–пиноцитоз, состоящий в том, что клеточная мембрана окружает внеклеточный
материал с образованием маленького пузырька. Образовавшийся пузырек погружается
в цитоплазму и мигрирует к базальной поверхности клетки, где заключенный в
пузырьке материал выбрасывается из энтероцита. По–видимому, пиноцитоз играет
определенную роль во всасывании интактных белков и других макромолекул.
Всасывание
воды. По всей длине
желудочно–кишечного тракта перенос воды в обоих направлениях–от слизистой
поверхности к серозной (всасывание) и от серозной к слизистой (секреция)–это
чисто пассивный процесс. Пока не совсем ясно, как в основном
осуществляется перенос воды–через клетки или через межклеточное пространство.
Известно, что объем трансэпителиального тока воды уменьшается параллельно
уменьшению осмотического градиента в направлении от верхних отделов кишечника к
нижним. Поскольку размер пор в плотных контактах также постепенно уменьшается в
том же направлении, очевидно, что межклеточный транспорт воды имеет более
важное значение. Движущей силой трансэпителиального переноса воды служат
гидростатический и, главное, осмотический градиенты, создаваемые активным
транспортом натрия между просветом кишечника и межклеточным пространством
(рис. 29.15). В результате активного выведения натрия из клетки через
базолатеральную мембрану межклеточная жидкость становится гипертонической.
Создающийся при этом устойчивый осмотический градиент вызывает ток воды
из просвета кишечника в межклеточное пространство через плотные контакты или
через эпителиальную клетку. Ток воды создает в межклеточном пространстве
градиент гидростатического давления. Это давление направлено как в сторону
слизистой, так и в сторону серозной поверхности, но, поскольку базальная
мембрана обладает значительно большей проницаемостью, чем плотные контакты,
вода стремится выйти из межклеточного пространства именно в этом направлении,
т.е. в сторону субэпителиальных капилляров. Таким образом, именно гидростатическое
давление в межклеточном пространстве служит движущей силой передвижения воды и
растворенных веществ в субэпителиальные капилляры, и это передвижение может
иметь место только в том случае, если гидростатическое давление в межклеточном
пространстве больше фильтрационного давления в капиллярах.
На рис. 29.15, Б приведена схема механизма
транспорта воды в виде модели с тремя компартментами. Компартменты
представляют собой просвет кишечника вблизи эпителия (1), межклеточное
пространство (2) и субэпителиальное пространство (3). Прежде всего в результате
активного транспорта натрия повышается осмотическая концентрация в компартменте
2, что вызывает вход воды по осмотическому градиенту через плотные контакты (с
серозной поверхности никакого поступления воды не происходит, так как
концентрационный градиент между компартментами 1 и 2 больше,
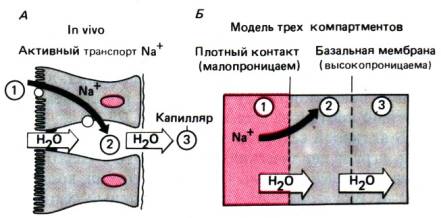
Рис. 29.15. Механизмы всасывания воды.
Поглощение воды полностью пассивный процесс, возникающий вследствие
функциональной асимметрии эпителия. Эта асимметрия обусловлена осмотическим
градиентом между просветом кишечника и межклеточным пространством, а также
гидростатическим давлением в межклеточном пространстве и разной проницаемостью
плотных контактов и базальной мембраны
чем между компартментами 2 и 3). По мере поступления
воды в компартмент 2 в нем увеличивается гидростатическое давление, под
действием которого вода перемещается в компартмент 3. Таким образом, весь
процесс основан на совместном действии активного и пассивного транспорта
в трех компартментах, разделенных мембранами, одна из которых (плотный контакт)
малопроницаема для ионов, а вторая (базальная мембрана) обладает высокой
проницаемостью. В тонком кишечнике, где в плотных контактах имеются
относительно крупные поры, интенсивное поступление воды из просвета кишечника
противостоит тенденции к гипертоничности межклеточной жидкости, и ионы Na+ всасываются в кровь в виде раствора, изотоничного плазме. В толстом
кишечнике
поры плотных контактов намного мельче, поэтому вода поступает из просвета кишечника
в межклеточное пространство значительно медленнее и лишь слегка разводит
межклеточную жидкость, в связи с чем всасываемый раствор является
гипертоническим по отношению к плазме, а жидкость каловых масс,
следовательно,– гипотонической.
Выделение минеральных солей и воды. Выделение веществ в просвет
кишечника наблюдается в тех случаях, когда от серозной поверхности к слизистой
переносится больше жидкости, чем в противоположном направлении. Такой процесс
может иметь место как в случае уменьшения тока жидкости от слизистой
поверхности к серозной (мальабсорбция), так и при увеличении переноса от
серозной поверхности к слизистой (рис. 29.6). Поскольку транспорт воды–это
полностью пассивный процесс, для превращения всасывания в секрецию должно
измениться направление осмотического или гидростатического градиента. Таким
образом, предпосылкой для секреции воды всегда служит секреция электролитов.
Существует несколько возможных механизмов секреции жидкости: 1) активная
секреция анионов; 2) уменьшение активного всасывания; 3) высокая осмолярность в
просвете кишечника (рис. 29.34); 4) повышение гидростатического давления на
серозной поверхности; 5) повышение проницаемости плотных контактов для ионов
(рис. 29.16).
В активной секреции
электролитов центральную роль играет циклический АМФ (цАМФ). Некоторые бактериальные
токсины (холерный токсин, колитоксин) и ряд гормонов
(простагландины, секретин, вазоактивный интестинальный полипептид) вызывают
повышение содержания цАМФ в энтероцитах. Изменяя концентрацию ионов Са2+
в клетках, цАМФ повышает проницаемость для С1– в криптах слизистой
поверхности энтероцитов. Хлорид выходит из клеток в просвет кишечника, а за ним
следуют ионы натрия и вода; первые–по принципу сохранения электронейтральности,
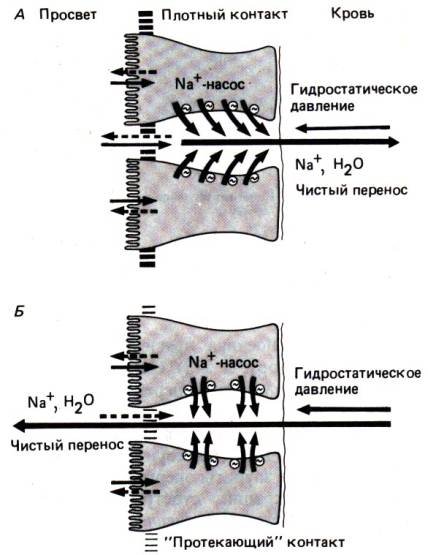
Рис. 29.16. Секреция воды и электролитов через межклеточное пространство. Дигидроксипроизводные желчных кислот и слабительные препараты,
вызывающие выведение воды, повышают проницаемость плотных контактов. Это
«протекание» ликвидирует функциональную асимметрию эпителия, и под действием
гидростатического давления на субэпителиальной стороне направление переноса
меняется на противоположное (по [38])
а вторая–под действием
осмотического давления. В других, указанных выше ситуациях секреция жидкости
осуществляется пассивным путем.
29.2. Ротовая полость, глотка и пищевод
Ротовая полость, глотка и
пищевод образуют функциональную единицу, назначение которой–предварительная
обработка пищи перед прохождением ее по желудочно–кишечному тракту. Здесь пища
подвергается измельчению и смачиванию слюной, после чего поступает в желудок.
Жевание. На этом подготовительном
этапе пища разрезается на куски и перетирается. Такое измельчение не абсолютно
необходимо для переваривания и всасывания, но сильно облегчает эти процессы. В
пережевывании пищи участвуют верхние и нижние челюсти, зубы, поперечнополосатая
мускулатура нижней челюсти, язык, щеки, дно ротовой полости и нёбо. Ритмичный
процесс жевания осуществляется в основном как непроизвольный рефлекторный акт.
При соприкосновении частиц пищи с нёбом и зубами
возникает рефлекторное жевательное движение, при котором пища
перемещается из одной стороны в другую, а также вперед и назад. Один такой цикл
длится 0,6–0,8 с. Прилагаемое при этом усилие составляет около 100–250 Н
в области резцов и 300–900 Н в области коренных зубов, достигая при
максимальном усилии 1500 Н. Чем дальше от зубов, тем меньше прилагаемое усилие:
так, на расстоянии 1 см оно составляет 400 Н, а на расстоянии 2 см–только 120 Н
[16]. Для максимального измельчения пищи необходим полный набор зубов, и
отсутствие нескольких из них нельзя компенсировать более интенсивным или
продолжительным жеванием.
При помощи языка пищевой комок удерживается между
челюстями в пределах жевательной поверхности зубов. Твердая пища измельчается
до частиц диаметром в несколько миллиметров. Благодаря слюноотделению,
стимулированному жеванием, пища приобретает консистенцию, необходимую для
проглатывания. Жевание и растворение или суспендирование твердых компонентов
пищи в слюне усиливают вкусовые ощущения, которые вызывают рефлекс,
стимулирующий слюноотделение и секрецию желудочного сока.
Секреция слюны
Слюна образуется в ротовой
полости со скоростью около 1 л в сутки. Она смачивает полость рта,
облегчая таким образом артикуляцию, смазывает пережеванную пищу и способствует
вкусовым ощущениям. Слюна имеет важное значение для сохранения зубов; в ее
отсутствие они поражаются кариесом и выпадают. Слюна очищает также полость рта
и обладает бактерицидным действием благодаря присутствию в ней лизоцима
и ионов тиоцианата. При уменьшении слюноотделения возникает чувство
жажды, стимулирующее потребление жидкости. Наконец, под действием слюны
начинается переваривание углеводов.
Слюнные
железы. Многочисленные мелкие
выделяющие слизь железы, расположенные в слизистых щек и языка, не обеспечивают
адекватного смачивания ротовой полости. Оно обеспечивается тремя парами крупных
желез– околоушных, подчелюстных и подъязычных. Каждая такая
железа состоит из концевых секреторных клеток и системы внутри– и междолевых
протоков. По гистологическому строению и характеру выделяемой слюны различают серозные
железы, вырабатывающие слюну, богатую белком, водой и минеральными солями,
(околоушные железы) и железы смешанного типа, слюна из которых содержит
еще и мукополисахариды (подчелюстные и подъязычные железы).
Нервная регуляция секреции. Слюноотделение находится
под контролем симпатической и парасимпатической нервных систем. Индуцируют
слюноотделение секреторные центры продолговатого мозга, получающие
афферентные сигналы из ротовой полости и нёба (вкусовые и тактильные), из
носовой полости (запахи) и из высших отделов мозга (представление о еде). Парасимпатическая
стимуляция вызывает образование больших количеств слюны с низким содержанием
белка. В то же время симпатическая стимуляция, которую можно
воспроизвести введением в шейную артерию норадреналина, вызывает секрецию
относительно небольшого количества вязкой слюны из подчелюстных и подъязычных
желез (но не из околоушных желез); при этом происходит также сужение кровеносных
сосудов и сокращение слюнных протоков.
Передача нервного
возбуждения на ацинозные клетки и их ответ (т.е. сопряжение стимула и
секреции) происходит по схеме, изображенной на рис. 29.3. Секреция
минеральных солей и воды слюнными железами стимулируется главным образом
адренергическими агентами и ацетилхолином, а секреция ферментов –
преимущественно при участии b–адренергических рецепторов.
В отсутствие стимуляции слюнные железы секретируют
слюну со скоростью около 0,5 мл/мин. При обезвоживании, испуге или стрессе
количество слюны уменьшается, а во время сна и наркоза слюноотделение почти
полностью прекращается. Обонятельные стимулы (например, запах амилонитрила)
могут вдвое повышать скорость слюноотделения, а при жевании она увеличивается в
2,5 раза. Крупные частицы пищи и вкусовые раздражители еще более усиливают
выделение слюны. Например, если взять в рот 0,5 М раствор лимонной кислоты,
скорость слюноотделения увеличится до 7,4 мл/мин.
По опыту все мы знаем, что
условные рефлексы, вызываемые видом пищи, звуками, сопровождающими ее прием,
или просто мыслями о ней, могут повысить слюноотделение. В состоянии покоя
вклад отдельных желез в общий объем выделяемой слюны распределяется следующим
образом: 71% выделяют подчелюстные железы, 25%–околоушные и 4% –подъязычные.
При стимуляции это соотношение изменяется и составляет 63, 34 и 3%
соответственно.
Скорость кровотока через слюнные железы в покое составляет
0,1–0,6 мл •мин–1 •г–1 ткани, а при стимуляции возрастает
приблизительно в 5 раз.
Состав слюны. Слюна состоит на 99% из
воды, поэтому ее удельный вес близок к 1,0. Наиболее важными минеральными
компонентами слюны являются Na+, К+ , С1–
и НСО3– . Первичный секрет, вырабатывающийся в дольках
слюнных желез, изотоничен крови. Но во время прохождения через протоки,
обладающие низкой проницаемостью для воды, слюна становится гипотонической,
поскольку реабсорбция Na+ сопровождается пассивным
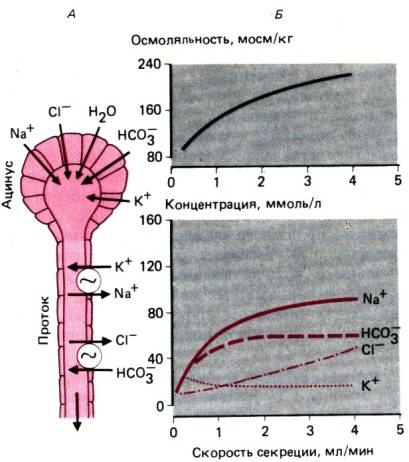
Рис. 29.17. Транспорт электролитов в
слюнных железах (А) и состав электролитов в слюне как функция скорости
секреции (6). Изотонический первичный секрет, выделяемый ацинусом, при
прохождении через протоки подвергается изменениям в результате поглощения или
секреции ионов и воды. Поскольку протоки относительно непроницаемы для воды,
слюна становится гипотонической. Наибольшее изменение состава слюны наблюдается
при низкой скорости секреции. При более высокой скорости секреции для обменных
процессов остается меньше времени и состав слюны сохраняется более близким к
составу первичного секрета (модифицированный рисунок из [1])
поглощением С1–. Конечная осмолярность
слюны составляет 2/3 величины осмолярности плазмы. Состав электролитов слюны
зависит от скорости секреции: при увеличении объема слюны концентрация Na+ и С1 повышается, а концентрация К+ падает (рис.
29.17), поскольку с увеличением скорости тока слюны остается меньше времени для
реабсорбции Na+ и секреции К+. В условиях покоя рН слюны
составляет 5,45–6,06, а при стимуляции повышается до 7,8.
Слюнные железы секретируют различные макромолекулы
– амилазу, гликопротеины, мукополисахариды, лизоцим, иммуноглобулины и
вещества, определяющие группу крови. Наибольшее функциональное значение имеет
a–амилаза, секретируемая
преимущественно околоушными железами, и мукопротеины, секретируемые
подчелюстными и подъязычными железами. a–Амилаза стабильна при рН
4–11 и максимально активна при рН 6,9. Она гидролизует a–1,4–гликозидную связь в полисахаридах,
расщепляя крахмал на мальтозу и мальтотриозу. Заболевания слюнных желез
(например, синдром Шегрена – одна из разновидностей ревматоидной
патологии) вызывают «ксеростомию», или сухость во рту, и это состояние
предрасполагает к образованию язвы желудка, кариесу зубов и затрудненности
жевания и глотания.
Глотание
Сформированный пищевой комок
проглатывается путем проталкивания через три отдела–ротовую полость, глотку и
пищевод. Таким образом, акт глотания включает три фазы–ротовую, глоточную и
пищеводную, из которых только первая имеет произвольный характер.
Поперечнополосатая мускулатура ротовой полости и глотки сокращается только под
влиянием нервных импульсов, поступающих из центральной нервной системы, а в их
отсутствие находится в расслабленном состоянии. Стенки двух верхних третей
пищевода состоят из. гладких мышц, поэтому находятся под автономным контролем.
Ротовая и глоточная фазы. В первой фазе акта глотания кончик языка
поднимается, отделяет часть пищевого комка и передвигает его к середине ротовой
полости, в промежуток между корнем языка и твердым нёбом (рис. 29.18, А).
Губы и челюсти смыкаются, мягкое нёбо поднимается, и передняя часть языка
проталкивает пищевой комок в верхнюю часть глотки (рис. 29.18, Б).
Мягкое нёбо и сократившиеся мышцы глотки образуют перегородку, которая отделяет
носоглотку от ротовой полости. При избирательном расстройстве этого смыкания
(что может иметь место при полиомиелите) пищевой комок проталкивается в
носоглотку и проглоченная жидкость вытекает через нос.
Когда язык проталкивает
пищевой комок еще дальше назад, дыхание на короткий момент рефлекторно
прерывается. Гортань поднимается и перекрывает вход в дыхательные пути
(рис. 29.18, В). Под давлением пищевого комка надгортанник изгибается
над входом в трахею, предотвращая забрасывание в нее кусочков пищи. При
нарушении этого механизма пища «идет не в то горло». Подталкиваемый мышцами
глотки и языком под давлением 4–10 мм рт.ст. пищевой комок проскальзывает над
надгортанником в пищевод (рис. 29.18, Г).
В это же время открывается верхний сфинктер пищевода (в основном благодаря
расслаблению перстнеглоточной мышцы) (рис. 29.18, Д).
Ротовая фаза носит, таким
образом, произвольный характер, поступление же пищевого комка в глотку вызывает
безусловный рефлекс за счет раздражения рецепторов ротовой полости и
глотки. Афферентные импульсы передаются по языкоглоточному нерву и
верхней гортанной ветви блуждающего нерва в двигательные нейроны,
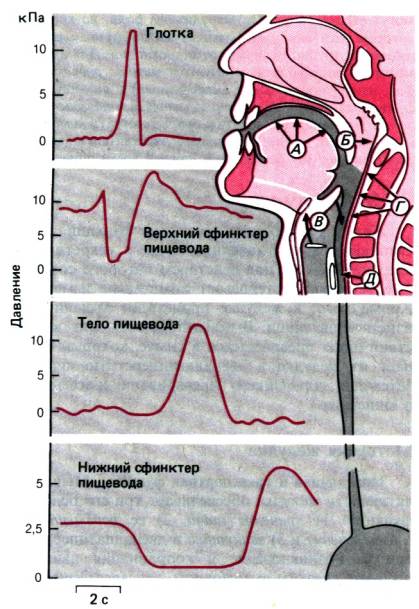
Рис. 29.18. Рото–глоточная и пищеводная
фазы глотания. А. Язык прижимается к твердому нёбу. Б. Носоглотка
перекрывается мягким нёбом. В. Гортань поднимается, и надгортанник
перекрывает вход в трахею. Г. Мышцы глотки начинают сокращаться. Д.
Рефлекторно открывается верхний сфинктер пищевода. Кривые (слева) показывают
изменения давления в глотке, верхнем сфинктере пищевода, теле пищевода и нижнем
сфинктере пищевода
иннервирующие глотку. Последние образуют пять
основных групп, локализованных в двигательных ядрах тройничного, лицевого и
подъязычного нервов, а также в двойном ядре блуждающего нерва и в сегментах
С1/СЗ спинного мозга. После стимуляции «центра глотания», находящегося в
продолговатом мозгу, сложный процесс глотания происходит непроизвольно, по
заданной программе. В норме взрослый человек совершает за сутки около 600
глотательных движений–200 во время еды, еще 350 в состоянии бодрствования и 50
во время сна. Тонкая регуляция этого сложного процесса требует обильной
иннервации и небольшого числа двигательных элементов. Соотношение числа нервных
и мышечных волокон составляет в глоточных мышцах 1:2–1:6, тогда как в
икроножной мышце оно равно 1:2000.
Пищеводная фаза. Пройдя через верхний
сфинктер пищевода, пищевой комок попадает в пищевод–мускулистую трубку длиной
25–35 см. Стенки пищевода состоят из наружного слоя продольных мышц и
внутреннего слоя циркулярных мышц. В пищеводе различают три отдела: 1) верхний
сфинктер пищевода–участок длиной 2–4 см, обладающий высоким тоническим
напряжением и находящийся под контролем нервов, вызывающих его расслабление при
глотании; 2) тело пищевода, находящееся под контролем внешних нейронов и
независимых эндогенных механизмов, и 3) нижний сфинктер пищевода –
вторая зона создающегося в просвете высокого давления, перекрывающая вход в
желудок.
Большая часть пищевода
расположена в грудной клетке, и внутреннее давление в нем на 4 5 мм рт. ст.
ниже атмосферного. В связи с этим в пищевод должен был бы проходить воздух,
если бы верхний сфинктер не перекрывал его верхний конец под давлением 50–100
мм рт.ст. Нижний сфинктер пищевода препятствует забрасыванию в пищевод
содержимого желудка благодаря тому, что развивающееся в нем давление на 15–25
мм рт.ст. выше, чем на дне желудка.
В верхней трети пищевода
мышечный слой состоит из поперечнополосатой мускулатуры, а в нижних двух
третях–из гладкой. Изнутри пищевод выстлан многослойным плоским эпителием,
содержащим немногочисленные слизистые железы. Узкая зубчатая линия границы
разделяет эпителий пищевода и цилиндрический эпителий желудка. Иннервируется
пищевод главным образом блуждающим нервом. Поперечнополосатые мышцы
верхнего отдела пищевода иннервируются соматическими волокнами, а гладкие мышцы
нижнего отдела имеют автономную иннервацию, типичную для желудочно–кишечного
тракта (рис. 29.2).
Активация продольных и циркулярных
мышц имеет разный характер. Продольные мышцы иннервируются холинергическими
волокнами и находятся в сокращенном состоянии до тех пор, пока действует
стимул. В циркулярных мышцах в ответ на нервный импульс сначала развивается
небольшое кратковременное напряжение, и только после выключения действия
стимула начинается сокращение. Вдоль пищевода существует градиент латентности,
и дистальные отделы отвечают на стимуляцию позже, чем проксимальные.
Ацетилхолинергические препараты на сокращения пищевода не действуют. В то же
время они подавляются нервным ядом тетродотоксином, поэтому предполагают, что
нейромедиатором в циркулярных мышцах служит какой–то желудочно–кишечный пептид,
скорее всего ВИП. Сокращения
пищевода, продолжающие акт глотания и возникающие в верхнем отделе пищевода,
называют первичной перистальтикой (рис. 29.18).
Вторичная перистальтика начинается с афферентных
импульсов, поступающих из самого пищевода,
например возникающих под действием давления,
создаваемого остатками пищевого комка, которые оказались непродвинутыми под
действием первичной перистальтики. Длина перистальтической волны пищевода
равна 2–4 см и распространяется со скоростью 2–4 см/с, достигая нижнего
сфинктера пищевода приблизительно через 9 с (рис. 29.18). Скорость
прохождения волны может колебаться в зависимости от консистенции пищевого
комка и положения тела. При вертикальном положении тела вода доходит до желудка
за 1 с, слизистая масса–за 5 с, а твердые частицы–за 9–10 с. По мере
прохождения перистальтической волны давление увеличивается и в нижнем отделе
пищевода достигает 30–120 мм рт.ст. Величина давления повышается с
увеличением размеров пищевого комка, и при «сухом» глотании (т. е. в отсутствие
пищевого комка) она значительно меньше. Нижний сфинктер пищевода открывается,
пропуская пищу в желудок, а затем вновь закрывается, т. е. после короткой фазы повышенного
давления восстанавливается тонус покоя. Это расслабление имеет
рефлекторную природу и осуществляется под действием блуждающего нерва.
Предполагают, что нейромедиатором в данном случае служит ВИП.
На давление в области нижнего сфинктера пищевода
влияют многие факторы. Оно повышается при увеличении давления в брюшной
полости, вызванного сокращениями брюшных мышц, при защелачивании желудочного
сока, а также после приема белковой пищи и уменьшается после приема таких
веществ, как жиры, шоколад, мятные лепешки, алкоголь или никотин. В регуляции
давления в области верхнего сфинктера пищевода участвуют нейрогенные (холинергические),
миогенные и гормональные факторы, при этом гастрин, мотилин,
вещество Р, панкреатический полипептид, вазопрессин и ангиотензин II повышают
давление, а секретин, холецистокинин, глюкагон, глюкозозависимый
инсулинотропный полипептид (ГИП) и ВИП снижают его. Давление уменьшается также
под действием прогестерона, и этим объясняется изжога, часто сопровождающая
беременность, во время которой уровень прогестерона повышен.
Патофизиологические аспекты. Нарушения нормальной
функции пищевода и его сфинктера могут вызывать определенные клинические
симптомы. При ахалазии нарушается координация перистальтики, и нижний
сфинктер пищевода не может при глотании расслабляться в достаточной степени. В
результате пища задерживается в пищеводе и он растягивается (мезофагия).
Причиной этого расстройства служат дегенеративные изменения в ауэрбаховом
сплетении. В Южной Америке оно известно как синдром Чага, вызываемый там
трипаносомами. Причина же его возникновения в Северном полушарии неизвестна.
Другой вид патологии, имеющий важное клиническое значение, возникает, когда
сфинктер закрывается неполностью, вследствие чего содержимое желудка забрасывается
в пищевод, что может вызвать повреждение слизистой вплоть до ее воспаления и
образования язв (рефлюкс–эзофагит). Беспорядочные, интенсивные
сокращения пищевода, называемые третичными сокращениями, могут вызывать острые
загрудинные боли, по характеру которых этот синдром, называемый диффузным
спазмом пищевода, трудно отличить от стенокардии. Для изучения моторики
пищевода используются методы радиокимографии, измерения давления (манометрия)
с помощью перфузионных катетеров, функциональной сцинтиографии после
заглатывания радиоактивного вещества и повторных измерений рН, когда с
помощью чувствительного зонда регистрируют рН рефлюкса в нижней трети пищевода.
29.3. Желудок
Желудок выполняет несколько
функций. В нем скапливается проглоченная пища и секретируется желудочный сок,
под действием которого содержимое желудка претерпевает химические изменения.
Кроме того, в желудке происходит механическое измельчение пищи. В результате
всех этих воздействий пища превращается в химус, который поступает из желудка в
двенадцатиперстную кишку и затем для дальнейшего переваривания и всасывания в
кишечник.
Моторика
желудка
Запасающая и транспортная функции. Мышечная активность желудка
обеспечивает три его основные функции: 1) хранение пищи, 2) ее перемешивание
и измельчение и 3) эвакуацию в двенадцатиперстную кишку.
Возникновение и координация движений желудка связаны в основном с непосредственным
ответом гладких мышц на действие целого ряда нейромедиаторов и гормонов при
участии рецепторов. Эти регуляторы модулируют возбудимость и сократительную
активность гладкомышечных клеток во время фазы активного переваривания в
желудке и между приемами пищи («голодная фаза»).
Мускулатура
желудка состоит из трех слоев:
наружного слоя продольных мышц, хорошо развитого среднего
слоя циркулярных мышц и внутреннего слоя косых мышц, наиболее
развитых в передней и задней стенках. Между мышечными слоями и под ними
находятся собственные нервные сплетения желудка–ауэрбахово и мейсснерово
(рис. 29.2). К этим сплетениям подходят холинергические и адренергические
нервные волокна, которые выделяют стимулирующие или ингибирующие медиаторы.
Проксимальный и дистальный отделы желудка сильно различаются по функциям и
характеру регуляции.
Желудок как резервуар. В проксимальном отделе желудка отсутствует
какой–либо ритм возбуждения и перистальтики. В нем поддерживается тонус,
зависящий от наполнения желудка. Иными словами, основное назначение
проксимального отдела желудка–хранение поступившей в него пищи. Еще до того,
как пищевой комок поступает из пищевода в желудок, давление в нем падает (рецептивное
расслабление). Благодаря рецепторам растяжения мышечный тонус желудка
изменяется таким образом, что его объем увеличивается без какого–либо повышения
давления (адаптивное расслабление). Этот процесс происходит под действием
ингибирующих волокон блуждающего нерва и гормонов; в частности, эластичность
желудка увеличивается под действием холецистокинина. При поступлении
порции пищи в желудок относительно твердые ее компоненты располагаются слоями,
а жидкость и желудочный сок обтекают их снаружи и поступают в дистальный отдел
желудка. Медленные тонические сокращения создают постоянное давление, под
действием которого пища постепенно перемещается в сторону привратника.
Перемещение в желудке твердого содержимого.
Тело желудка в отличие от его дна содержит миогенные
пейсмекеры, которые генерируют трехфазные циклические потенциалы –
медленные волны, начинающиеся в области большой кривизны и захватывающие
антральный отдел (рис. 29.19). Эти волны распространяются в сторону пилорического
отдела с частотой 3/мин и ответственны за перемещение пищи в желудке. Медленные
волны вызывают сокращения, но при этом не каждая волна связана с сокращением.
Медленная волна создает в мышцах потенциал, близкий к порогу активации, а после
этого нервные и гормональные воздействия индуцируют сокращение и определяют его
силу и продолжительность.
Сильные круговые перистальтические волны в
дистальном отделе желудка проталкивают его содержимое в сторону привратника и
двенадцатиперстной кишки. Жидкость быстро эвакуируется в двенадцатиперстную
кишку, и ее объем в желудке экспоненциально уменьшается. Твердые компоненты
пищи не проходят через привратник до тех пор, пока не будут измельчены
до частиц размером не более 2–3 мм; 90% частиц, покидающих желудок, имеют в
диаметре не более 0,25 мм. Когда перистальтические волны достигают дистального
участка антрального отдела, привратник сокращается. Пилорический отдел,
образующий самую узкую часть желудка (но не являющийся сфинктером в собственном
смысле этого слова) в месте его соединения с двенадцатиперстной кишкой,
закрывается еще до того, как антральный отдел полностью отгораживается от тела
желудка. Пища под давлением перемещается обратно в желудок, при этом твердые
частицы трутся одна о другую и еще больше измельчаются.
Опорожнение желудка регулируется вегетативной
нервной системой, интрамуральными нервными сплетениями и гормонами. В
отсутствие импульсов от блуждающего нерва (например, при его перерезке)
перистальтика желудка значительно ослабевает и опорожнение желудка замедляется.
Перистальтика желудка усиливается под действием таких гормонов, как
холецистокинин и, особенно, гастрин, и подавляется секретином, глюкагоном, ВИП
и соматостатином. Скорость опорожнения желудка –Е (уменьшение объема в единицу
времени)–зависит от разности давления в желудке (Рж) и в двенадцатиперстной
кишке (Рд) и от резистентности пилорического отдела (Rп)
Е=
(Рж–Рд)/Rп (2)
Благодаря свободному
прохождению жидкости через привратник скорость ее эвакуации зависит главным образом
от разности давлений Рж–Рд, причем основным регулятором служит давление
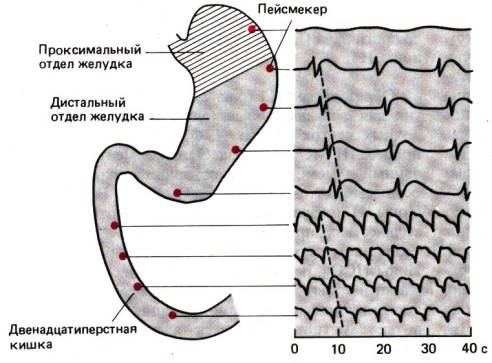
Рис. 29.19. Миоэлектрические потенциалы (медленные волны) в
желудке и двенадцатиперстной кишке. Тоническое сокращение проксимального отдела
желудка поддерживается без участия медленных волн. Медленные волны возникают в
пейсмекере с частотой 3–4/мин; по мере их распространения вниз происходит сдвиг
фаз, видимый на кривых. В двенадцатиперстной кишке частота медленных волн равна
12/мин, и по мере продвижения в дистальному отделу также наблюдается сдвиг фаз.
Мышечные сокращения возникают при наложении слабых генераторных потенциалов с
образованием медленных волн, дающих начало потенциалам действия
в проксимальном отделе желудка. В то же время эвакуация
из желудка твердых частиц пищи зависит главным образом от резистентности
привратника (Rп), а следовательно, от размеров частиц. В регуляции
опорожнения желудка помимо его наполнения, размеров частиц и вязкости
содержимого играют роль рецепторы тонкого кишечника. Кислое содержимое
эвакуируется из желудка медленнее, чем нейтральное, гиперосмолярное
содержимое–медленнее, чем гипоосмолярное, а липиды (особенно содержащие жирные
кислоты с цепями из более чем 14 углеродных атомов) – медленнее, чем продукты
расщепления белков (кроме триптофана). В регуляции эвакуации участвуют как
нервные, так и гормональные механизмы, причем в ее угнетении особенно важную
роль играет секретин.
Крупные твердые частицы не могут выводиться из
желудка во время фазы пищеварительного опорожнения. Такие неперевариваемые
частицы с диаметром более 3 мм могут проходить через привратник только в
голодную фазу при участии специального механизма миоэлектрического комплекса. В
фазе 3 этого процесса происходят интенсивные сокращения дистального отдела
желудка, благодаря чему привратник, закрытый во время пищеварительной фазы,
остается открытым и крупные частицы могут проходить в двенадцатиперстную кишку.
Желудочная секреция
Клетки желудочных желез
секретируют в сутки 2–3 л желудочного сока, содержащего ионы и макромолекулы.
Кроме того, желудок секретирует в кровь гормон гастрин. Желудочную
секрецию можно разделить на базальную (в голодную фазу) и стимулированную
(пищеварительную). Секретируемая желудком кислота вызывает денатурацию
белков и активирует пепсиноген, превращая его в пепсин, который гидролизует
белки и обладает бактерицидным действием. Выделяемая желудком слизь
обволакивает химус и защищает слизистую. Наконец, в желудочном соке
присутствует внутренний фактор, необходимый для всасывания витамина В12.
Через слизистую желудка всасываются лишь немногие вещества, в том числе
некоторые ионы, например Na+, и жирорастворимые
вещества, например алкоголь, большие количества которых могут быстро
всасываться в желудке.
Функциональная анатомия слизистой желудка.
Полость желудка выстлана слизистой оболочкой,
секретирующей слизь, пепсиноген II, а также бикарбонат и натрий. Этот
цилиндрический эпителий содержит аргентаффинные клетки (к настоящему времени их
идентифицировано 9 видов), секретирующие гормоны эндокринным (непосредственно в
кровь), паракринным (от клетки к клетке) и нейрокринным (из нервных окончаний)
путем. Слизистая оболочка образует желудочные ямки, на дне которых открываются желудочные
железы. В каждом отделе желудка имеются железы определенного типа.
Различают три отдела.
1. Кардиальный отдел, представляющий собой узкое,
шириной 1–4 см, кольцо ниже отверстия пищевода, содержит трубчатые железы с
многочисленными извитыми ответвлениями.
2. Дно и тело желудка, составляющие в сумме 3/4
всего желудка, содержат прямые или слегка изогнутые железы, в стенках которых
помимо аргентаффинных и выделяющих слизь клеток присутствуют париетальные
(обкладочные) клетки, секретирующие кислоту, и главные
(зимогенные) клетки, секретирующие пепсиноген I и II.
3. Пилорический отдел, составляющий 15–20%
желудка, содержит просто разветвленные трубчатые железы, секретирующие в
основном слизь. Особенность этого отдела состоит в наличии G–клеток, вырабатывающих
гастрин.
Клетки, секретирующие слизь
и пепсиноген, сходны с клетками такого же типа из других отделов
желудочно–кишечного тракта. В то же время обкладочные клетки обладают
уникальной способностью секретировать сильно концентрированную соляную кислоту;
концентрация ионов Н+ в их секрете в миллион или более раз выше, чем
в крови. Эти клетки содержат многочисленные крупные митохондрии и характерные
внутриклеточные канальцы, выстланные множеством микроворсинок и
открывающиеся на апикальной поверхности клеток в просвет желез (рис. 29.20). В
течение 10 мин после стимуляции клетки претерпевают выраженные морфологические
изменения. Содержание в их цитоплазме тубуловезикулярных структур,
преобладающих в покое, уменьшается почти на 90%, а диаметр микроворсинок в
канальцах и отверстий протоков увеличивается в 4–6 раз. В этих микроворсинках
локализована (Н +–К + )–АТФаза, транспортирующая ионы Н+.
Образование
НС1 (рис. 29.21). Источником
энергии для активного транспорта протонов из обкладочных клеток служит АТФ,
а катализирует этот процесс H+–K+–AТФaзa. Этот
фермент, локализованный в мембранах секреторных микроворсинок, осуществляет
эквивалентный обмен H+ на K+. Ионы Н+
образуются в эквивалентных с НСО3– количествах при
диссоциации Н2СО3 и в очень небольшом количестве–при
диссоциации Н2О. Ионы НСО3– перемещаются по
градиенту концентрации в кровь в обмен на С1–. Когда этот процесс
достигает максимума, концентрация НСО3 в сыворотке заметно
повышается явление, получившее название «щелочного периода». Одновременно с
Рис. 29.20. Обкладочная клетка в покое (А)
и при стимуляции (Б). Я ядро, Р–гладкий эндоплазматический ретикулум,
М–митохондрия, К–внутри клеточный каналец, открывающийся в просвет железы, В
вакуоль. При стимуляции канальцы расширяются, а вакуоли увеличиваются в
размерах
ионами H+ в просвет желудка против
градиента концентрации и электрического градиента путем активного транспорта
выводятся ионы С1–, причем в секреторной фазе ионов С1–
секретируется больше, чем ионов Н+. В состоянии покоя секреция ионов
С1– в сочетании с активным поглощением ионов Na+ служит основным механизмом, определяющим разность
электрических потенциалов величиной 40–60 мВ между внутренней областью
желудка (отрицательный заряд) и серозной поверхностью (положительный заряд).
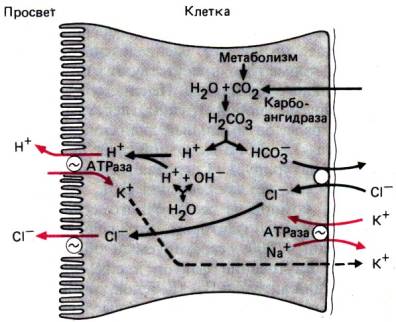
Рис. 29.21. Секреция HCI обкладочной клеткой. Ионы
Н+ переносятся в просвет при участии (Н+–К+–АТФазы,
встроенной в мембрану щеточной каемки. Ионы Сl– также активно переносятся в
просвет, а в клетку поступают в обмен на ионы НCO3–; ионы
Н+ образуются из Н2CO3 и в меньшей степени–из
воды [1]
Эта разность потенциалов свидетельствует о
целостности мембраны. При нарушении мембранного барьера, например под
действием желчных кислот, лизолецитина, экзогенного салицилата или алкоголя,
происходит обратная диффузия ионов Н+ в клетки из просвета желудка,
а ионов Na+ и белков – в просвет желудка, и уменьшается
разность потенциалов.
Образование бикарбоната в клетках слизистой.
Помимо кислоты слизистая желудка выделяет щелочной
секрет, образующийся в локализованных на ее поверхности и в железах слизистых
клетках. Этот процесс включает активную секрецию бикарбоната, причем
транспорт его электронейтрален и происходит, по–видимому, путем обмена ионов
бикарбоната на ионы С1–. В секреторной фазе образование Н+
в париетальных клетках во много раз превышает выделение бикарбоната клетками
слизистой, поэтому секрецию последнего трудно обнаружить НСО3–
нейтрализуется эквимолярными количествами Н+ с образованием СО2
и Н2О. Чтобы обнаружить секрецию НСО3–
необходимо специально блокировать выделение Н+. С этой целью можно
использовать антагонисты рецепторов гистамина (блокаторы Н–2–рецептора),
поскольку гистамин стимулирует секрецию H+.
Вместе с желудочной слизью бикарбонат защищает
слизистую от повреждающего действия желудочного сока. За счет секреции HCI
желудочными железами значение рН в просвете ямок и полости желудка намного
ниже, чем на поверхности выстилающих их слизистых и цилиндрических клеток..
Выделяемый этими клетками НСО3– задерживается в
прилежащем неперемешивающемся слое жидкости или слизи, что обеспечивает поддержание
разности рН, который в просвете желудка равен приблизительно 2,0, а на
поверхности клеток–около 7,0. На поверхность слизистых клеток желез поступают
также дополнительные количества ионов НСО3–, образованные
в обкладочных клетках в фазе активной секреции (рис. 29.21). Последние выделяют
ионы НСО3– в кровь, из которой те поступают на
поверхность слизистой через образуемые капиллярами петли, перпендикулярные
просвету желудка. Через капилляры кроме того удаляется НС1, диффундирующая в
них из просвета желудка. Кровоток в этих капиллярах регулируется простагландином
Е2. Таким образом, простагландин Е2 наряду с
бикарбонатом и неперемешивающимся слоем играет важную роль в защите слизистой
желудка.
Недавно появились сообщения о
веществах, которые блокируют образование НС1 за счет угнетения (H+–K+)–АТФазы. Эти вещества
представляют собой производные бензимидазола (например, омепрозол). При
однократном введении человеку они ингибировали выделение кислоты в течение 48
ч, а в условиях in vitro подавляли стимулированное
цАМФ и К+ образование кислоты даже в
тех случаях, когда были неэффективны блокаторы Н2–рецепторов и атропин [21].
Полученные данные свидетельствуют о том, что активация (Н+–К+–АТФазы–это относительно позднее,
а возможно, и последнее звено в цепи событий, приводящих к секреции кислоты.
Состав желудочного сока. Содержание в желудочном соке электролитов
зависит от скорости секреции. Слизистые клетки постоянно секретируют Na+, К+, С1–
и НСО3– а обкладочные клетки при стимуляции
выделяют H+ и Сl-. С повышением скорости
секреции доля выделений обкладочных клеток в общем секрете увеличивается; при
этом в нем снижается содержание Na+ и полностью исчезает НСО3–
(рис. 29.22).
Важный компонент желудочного
сока–слизь. Она покрывает всю внутреннюю поверхность желудка, образуя слой
толщиной около 0,6 мм, который обволакивает слизистую и защищает ее от
механического и химического повреждения. Слизистый слой сохраняется постоянно и
при механическом его удалении или пептическом переваривании образуется вновь.
Основным компонентом слизи служит гликопротеин с мол. массой 2 млн. Это
полимер, состоящий из белковой основы и многочисленных углеводных боковых
цепей. Состав его концевой углеводной последовательности обнаруживает индивидуальные
генетически детерминированные различия, коррелирующие с иммунологическими
группами крови системы АВО.
Другой важный компонент
желудочного сока пепсиноген, представляющий собой смесь предшественников
протеаз. Под действием кислоты пепсиноген превращается в активный
протеолитический фермент пепсин, причем, начавшись, это
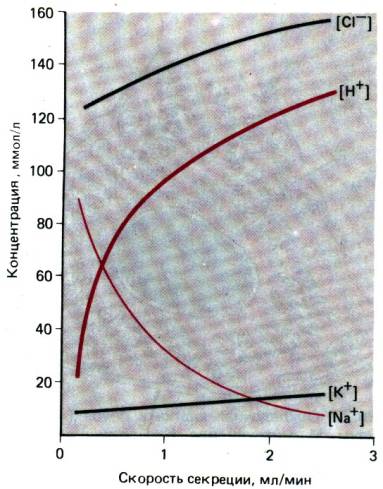
Рис. 29.22. Состав электролитов
желудочного сока как функция скорости секреции. В состоянии покоя преобладает
секреция слизистых клеток, расположенных на поверхности и в желудочных ямках.
По мере увеличения скорости секреции концентрация ионов H+ повышается, а ионов Na+ уменьшается (по [11])
превращение идет далее по
принципу автокатализа. С помощью электрофореза обнаружено по меньшей мере 8
протеолитических ферментов. Пять быстро мигрирующих пепсиногенов, составляющих группу
I, обнаруживаются только в области главных и обкладочных клеток. Остальные
пепсиногены, составляющие группу II, распределены по всему желудку и,
кроме того, присутствуют в бруннеровых железах тонкого кишечника. Пепсин,
образующийся из пепсиногена любой группы, действует только в кислой среде;
оптимальный диапазон рН для его активности–1,8–3,5. В щелочной среде пепсин
необратимо денатурирует. Секреция пепсиногенов регулируется так же, как
секреция НС1.
Третий компонент желудочного сока, называемый
внутренним фактором,–это гликопротеин с мол. массой 42000, секретируемый
обкладочными клетками. Внутренний фактор и еще один белок, связывающий витамин
В12 (R–белок, от англ. rapid–быстрый, названный так в
связи с высокой подвижностью при электрофорезе), необходимы для всасывания
витамина В12. В кислом желудочном соке витамин В12 связывается главным
образом с R–белком, но в верхнем отделе тонкого кишечника под действием
ферментов панкреатического сока этот комплекс расщепляется и высвобождающийся
витамин В12 взаимодействует с внутренним фактором, образуя новый комплекс,
устойчивый к протеолизу и не всасывающийся в верхнем отделе тонкого кишечника.
Этот комплекс взаимодействует со специфическими рецепторами в тощей кишке,
после чего витамин В12 всасывается в кровь воротной вены. Часть поглощенного
витамина В12 депонируется в печени, а остаток в виде комплекса с белком транскобаламином
II циркулирует с кровью.
Регуляция желудочной секреции
Секреция желудочного сока натощак
составляет у человека 10% того количества, которое секретируется при
максимальной стимуляции. Перерезка блуждающего нерва или удаление антрального
отдела желудка, содержащего G–клетки, приводит к полному прекращению базальной
секреции, из чего следует, что она стимулируется гастрином и зависит от
фонового тонуса блуждающего нерва. Адекватным стимулом для секреции служит прием
пищи. Воздействие пищи начинается еще до ее приема и продолжается после
того, как он закончится. Процесс секреции включает цефалическую, желудочную и
кишечную фазы, которые до некоторой степени перекрываются.
Цефалическая
фаза. Начало этой фазы связано с
ожиданием пищи, представлением о ней, ее видом или запахом и, наконец,
вкусовыми ощущениями. Возникающие соответственно в разных участках мозга
нервные импульсы передаются в желудок по блуждающему нерву. При
перерезке блуждающего нерва цефалическая фаза исчезает. Предполагают, что
секрецию вызывает гастрин, высвобождающийся под действием блуждающего нерва,
поскольку денервация антрального отдела желудка практически блокирует секрецию.
Скорость секреции в цефалической фазе составляет 40–50% максимальной.
Эта взаимосвязь была подробно изучена И. П. Павловым
(1889 г.) (рис. 29.23). Он проводил опыты на эзофаготомированных собаках с фистулой
пищевода и изолированным желудочком, выкроенным из тела и дна желудка. Такой
желудочек представляет собой «карман» из участка слизистой оболочки,
открывающийся наружу и сохраняющий интактное кровоснабжение и иннервацию
волокнами блуждающего нерва. В опытах на подобных животных были получены
следующие результаты. 1, «Мнимое кормление», при котором проглоченная пища
выпадает из перерезанного отрезка пищевода, приводит к обильному выделению
желудочного сока в изолированном желудочке (цефалическая фаза; безусловнорефлекторный
компонент). 2. Если кормлению всегда предшествует звук колокольчика, то через
несколько дней один лишь этот звук приводит к выделению слюны и желудочного
сока из соответствующих фистул (цефалическая фаза; условнорефлекторный компонент).
Эти опыты положили начало изучению условных рефлексов.
Желудочная
фаза. Желудочную фазу секреции
вызывает растяжение желудка поступающей в него пищей и химическое воздействие
определенных компонентов пищи. Стимулы, связанные с растяжением желудка,
передаются по нервным путям, причем и афферентные, и эфферентные сигналы – по блуждающему
нерву и посредством местных интрамуралъных рефлексов. Химическая
стимуляция осуществляется в основном посредством выделения гастрина
G–клетками антрального отдела. Известно несколько гастринов. Гастрины G–17 I и
II, состоящие из 17 аминокислотных остатков, представляют собой физиологические
формы и обладают наибольшей активностью у человека. Гастрин G–34 состоит
из 34 аминокислотных остатков и имеет более продолжительный период
существования, но в 6 раз меньшую активность, чем G–17. Основными химическими
стимуляторами секреции в желудочной фазе служат продукты переваривания
белков –пептиды, олигопептиды и аминокислоты, особенно
Рис. 29.23. Опыт Павлова, демонстрирующий влияние центральной
нервной системы на секрецию желудочного сока у собак с фистулами пищевода и
желудка. А. Возникновение цефалической фазы секреции при виде пищи (безусловный
рефлекс). 6. Индукция секреции при звуке колокольчика, с которым у собаки
ассоциируется кормление (условный рефлекс)
триптофан и фенилаланин, а также кальций и
другие вещества, в том числе ионы Мg2+, алкоголь и кофеин.
Кишечная
фаза. Желудочную секрецию
стимулируют также растяжение тонкого кишечника и присутствующие в нем продукты
переваривания белков. Механизм стимуляции имеет главным образом гормональную
природу, при этом активным гормоном считается энтерооксинтин.
В регуляции желудочной секреции важную роль играют,
кроме того, ингибиторные воздействия. Кислая среда в антральном
отделе (рН ниже 3,0) угнетает секрецию гастрина в желудочной фазе. Существенный
ингибиторный эффект оказывает тонкий кишечник. Здесь механизм угнетения индуцируют
кислоты, жиры и гипертонические растворы, под действием которых
слизистая кишечника секретирует гормоны секретин и бульбогастрон, подавляющие
секрецию желудка. Жиры угнетают секрецию желудочного сока только после их
гидролиза, причем наиболее сильным действием обладают свободные жирные кислоты
с длиной цепи более десяти углеродных атомов. В этом механизме участвуют,
очевидно, нейроны, причем медиаторами служат нейротензин и ГИП.
Рецепторы секреторных клеток. К числу медиаторов, играющих роль первых посредников
в индукции секреции, относятся ацетилхолин, гистамин и гастрин. При
взаимодействии гормонов или эффекторных веществ со специфическими рецепторами,
локализованными на поверхности клеток, образуется второй посредник–цАМФ или
ионы Са+ (кальмодулин), индуцирующий специфическую активность
клетки. Предполагают, что
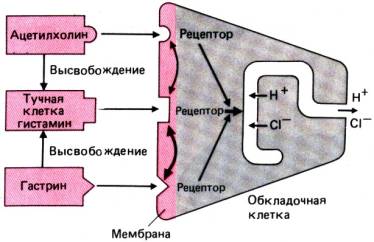
Рис. 29.24. Трехрецепторная
модель обкладочной клетки.
Предполагается, что существует сильное обязательное взаимодействие между
рецепторами гистамина и гастрина и более слабое, необязательное,–между
рецепторами гистамина и ацетилхолина. Антагонисты (H2–рецепторов блокируют
рецепторы гистамина (по [28])
обкладочные клетки имеют три вида рецепторов (рис.
29.24)–мускариновые рецепторы ацетилхолина (а не никотиновые, как в
других клетках), Н2–рецепторы гистамина (а не Н+–рецепторы,
как в других клетках) и рецепторы гастрина. Предполагают также, что существует
сильное обязательное взаимодействие между Н2–рецепторами и рецепторами
ацетилхолина. Наличие таких взаимодействий позволяет понять, почему блокаторы
Н +–рецепторов одновременно угнетают секрецию, вызываемую
гастрином или ацетилхолином. Применение конкурентных блокаторов Н2–рецепторов
(циметидин, ренитидин, феймотидин) и специфических блокаторов мускариновых
рецепторов в качестве ингибиторов секреции кислоты имеет большое значение при
лечении язвы.
Секреторная активность желудка. Через зонд, введенный в нижний отдел желудка, можно
отобрать желудочный сок для количественного анализа. Легко измеримым параметром
секреции является образование кислоты. Базальная секреция кислоты
составляет 2–3 ммоля Н+ в час, а при наличии опухоли, секретирующей
гастрин, она повышается в 10–20 раз. Максимальная скорость секреции (например,
при введении 6 мкг пентагастрина–синтетического аналога гастрина) на 1 кг веса
составляет 10–35 ммоль Н+ в час. У женщин эта величина несколько
меньше, чем у мужчин. У больных с язвой двенадцатиперстной кишки среднее
значение выше, чем у здоровых людей, однако существуют большие индивидуальные
различия. Раньше величина секреции кислоты служила важным показателем при
диагностике язвы, но, поскольку имеет место существенное перекрывание его
значений у здоровых и больных, этот показатель в настоящее время не используют.
Патофизиологические аспекты. Основные нарушения функции желудка связаны с
нарушением секреции. Уменьшение или отсутствие секреции не представляет
серьезной проблемы. Более чем у 50% людей старше 60 лет секреция кислоты
уменьшается в связи с прогрессирующей атрофией слизистой желудка, но и в
этом случае секреция поджелудочной железы обеспечивает адекватное переваривание
белков. Только при полном отсутствии внутреннего фактора прекращается
всасывание витамина B12, в результате чего нарушается
образование эритроцитов (эритропоэз) и развивается перннцнозная анемия.
При избыточной секреции соляной кислоты или нарушении
защитных механизмов может начаться самопереваривание слизистой желудка или
двенадцатиперстной кишки и образование пептических язв. Язвы могут также
возникать при гиперсекреции гастрина, связанной с разными причинами, в том
числе с некоторыми опухолями поджелудочной железы (опухолями островков, но не
бета–клеток), синдромом Золлингера – Эллисона или гиперплазией
G–клеток антрального отдела. При резекции желудка по методу Бильрота II
удаляют антральный отдел, зашивают верхний конец двенадцатиперстной кишки и
формируют анастомоз между желудком и тощей кишкой. Если при неправильном
проведении операции антральный отдел остается связанным с зашитой
двенадцатиперстной кишкой, то G–клетки, постоянно стимулируемые щелочным
содержимым кишечника, секретируют гастрин, который вызывает секрецию кислоты в
культе желудка.
Даже в тех случаях, когда .причина возникновения язвы
неясна (как чаще всего и бывает), можно ускорить заживление путем нейтрализации
желудочного сока или подавления его секреции. Поддержание значения рН на
уровне выше оптимального для действия пепсина позволяет блокировать один из
наиболее агрессивных компонентов желудочного сока, поэтому для лечения язв
используют противокислотные препараты, блокаторы Н+–рецепторов и
антихолинергетики.
29.4.
Поджелудочная железа
После эвакуации из желудка в
тонкий кишечник пища подвергается интенсивному перевариванию, и решающую роль в
этом процессе играет секреция поджелудочной железы, желчного пузыря и самого
тонкого кишечника. Наиболее важные компоненты панкреатического сока–это бикарбонат,
нейтрализующий кислый химус, и пищеварительные ферменты, расщепляющие основные
вещества в составе пищи. Секрецию сока поджелудочной железы регулируют в
основном гормоны секретин и холецистокинин (ХЦК), а также блуждающий нерв.
Поджелудочная железа, имеющая массу около 110 г,
способна выделять в сутки 1,5л секрета. Главный проток поджелудочной железы (вирсунгов
проток) проходит через всю железу и открывается в двенадцатиперстную кишку
позади общего желчного протока, а у 30–40% людей вместе с ним на верхушке
большого дуоденального (фатерова) сосочка.
Электролиты панкреатического сока. Панкреатический сок содержит
ряд электролитов и солей. Основные анионы в его составе–Сl– и НСО3–,
катионы –Na+ и K+. В отличие от слюны
панкреатический сок изотоничен плазме крови независимо от степени стимуляции.
Концентрация катионов при стимуляции остается постоянной, а концентрации С1–
и НСО3– меняются в противоположных направлениях (рис.
29.25). При максимальной секреции концентрация НСО3– составляет
13O–140 ммоль/л, а рН равен 8,2. Существуют две гипотезы, объясняющие
механизм разнонаправленного изменения концентраций двух катионов в
панкреатическом соке при изменении скорости секреции. Согласно гипотезе
обмена ионов НСО3– образуется в ацинусах в более
высокой концентрации, но в процессе прохождения секрета через протоки
обменивается на С1–; при высокой скорости секреции
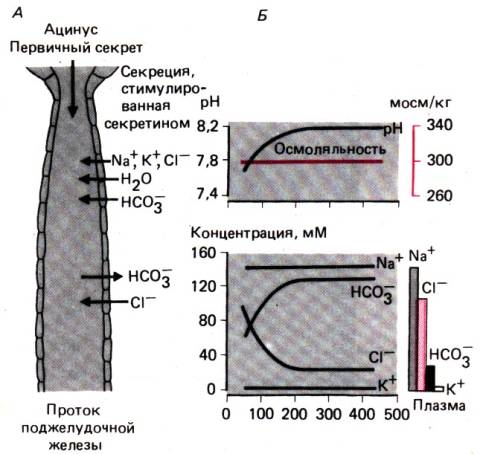
Рис. 29.25. Изменения первичного секрета
в протоке поджелудочной железы (А) и зависимость состава
панкреатического сока от скорости секреции (Б). При стимуляции
поджелудочной железы концентрация ионов Na+ и осмолярность остаются постоянными,
тогда как концентрации НСО3– и Сl– меняются во
взаимопротивоположных направлениях. Это изменение происходит вследствие
повышения скорости секреции ионов НСО3– в системе
протоков и уменьшения времени, в течение которого может происходить обмен ионов
НСО3– на ионы Сl–
время обмена сокращается.
Согласно гипотезе двух компонентов, ацинозные клетки выделяют ионы С1–
и Na+ в таких же концентрациях, в каких те содержатся в
плазме, тогда как клетки, выстилающие протоки, активно секретируют НСО3–,
поэтому при стимуляции последний преобладает. В настоящее время считают, что
действуют оба механизма. Помимо указанных выше ионов панкреатический сок
содержит Са2+, Mg2+ и Zn2+, а также
сульфат и фосфат.
Высокая концентрация НСО3– в
панкреатическом соке указывает на активный секреторный– процесс. Ионы Н +
активно переносятся из просвета протока в клетки в обмен на ионы Na +, которые выделяются в
просвет (рис. 29.26). Через базальную и латеральную поверхности клеток ионы Н4'
выходят из клеток в плазму (тоже в обмен на Na+ и реагируют здесь с НСО3–
с образованием СO2 и Н2О. СO2
диффундирует обратно в клетку и вместе с СO2 , образующимся в процессе
обмена веществ, выделяется в просвет панкреатических протоков. Здесь
карбоангидраза катализирует реакцию гидратации СO2 с образованием НСО3– .
Ферменты панкреатического сока. 90% белков панкреатического сока составляют
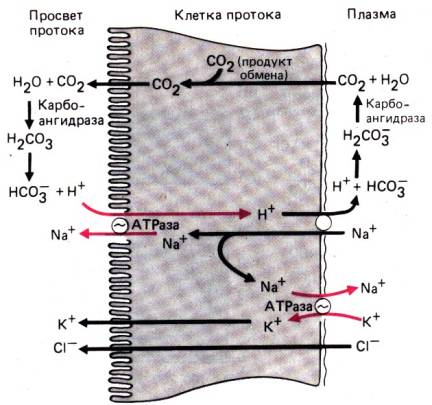
Рис. 29.26. Механизм секреции электролитов клетками
поджелудочной железы. Ионы H+ активно переносятся в обмен
на ионы Na+ из просвета протока в клетку, а затем в плазму, где
они взаимодействуют с ионами НСО3–с образованием Н2О
и СO2 .
СO2 диффундирует в просвет протока, где из него
образуются ионы НСО3– (по [1])
пищеварительные ферменты,
главным образом гидролазы, расщепляющие различные субстраты (табл. 29.2). Среди
них преобладают протеолитические ферменты–пептидазы. Пептидазы и фосфолипаза
А секретируются в виде зимогенов, т.е. предшественников, подлежащих активации,
тогда как липаза, амилаза и рибонуклеаза – в активной форме.
Активацию катализирует энтерокиназа–эндопептидаза, выделяемая слизистой
двенадцатиперстной кишки. Энтерокиназа катализирует превращение трипсиногена в трипсин,
после образования которого процесс продолжается уже путем автокатализа. Кроме
того, трипсин активирует, и другие протеазы. В панкреатическом соке
присутствует также ингибитор трипсина, который эффективно блокирует действие
трипсина при прохождении последнего через поджелудочную железу и таким образом
препятствует ее самоперевариванию.
В патологических условиях, однако, может происходить
самопереваривание поджелудочной железы собственными ферментами, приводящее к
возникновению синдрома, называемого острым панкреатитом. В наиболее тяжелых
случаях поджелудочная железа полностью разрушается и заболевание приводит к
смерти.
Находящиеся в ацинозных клетках зимогеновые гранулы
содержат все ферменты, присутствующие в секрете, в постоянном соотношении,
так что в панкреатическом соке оно также постоянно. Если рацион включает очень
большие количества какого– либо компонента, например жиров, возможны некоторые
изменения в соотношении ферментов, но для такой адаптации (в данном случае для
увеличения относительного количества липазы) требуется несколько недель.
Таблица 29.2. Ферменты, секретируемые поджелудочной железой
|
Характер действия
|
Участок гидролитического расщепления
|
|
Протеолитические
|
|
Эндопептидазы
|
Внутренние пептидные связи между соседними
аминокислотными остатками
|
|
Трипсин
|
Между
|
остатками основных аминокислот
|
|
Химотрипсин
|
остатками ароматических аминокислот
|
|
Эластаза
|
остатками гидрофобных аминокислот в эластине
|
|
Экзопептидазы
|
Концевые пептидные связи
|
|
Карбоксипептидазы А и В
|
СООН–конец (А – неосновные аминокислоты, В –
основные аминокислоты)
|
|
Аминопептидазы
|
N–конец
|
|
Амилолитические
|
a–1,4–Гликозидные связи в
полимерах глюкозы
|
|
a–амилаза
|
Эфирные связи
|
|
Липолитические
|
|
Липаза
|
В положениях 1 и 3 триглицеридов
|
|
Фосфолипаза А2
|
В положении 2 фосфоглицеридов
|
|
Холестеролаза
|
В эфирах холестерола
|
|
Нуклеолитические
|
|
Рибонуклеаза
|
Фосфодиэфирные связи между нуклеотидами в
рибонуклеиновых кислотах
|
Регуляция
панкреатической секреции
Гормональная и нервная
активация. Наиболее
эффективными стимуляторами экзокринного отдела поджелудочной железы служат
гормоны секретин и холецистокинин (ХЦК). Секретин стимулирует клетки,
выстилающие протоки и секретирующие главным образом бикарбонат, другие ионы и
воду. Холецистокинин стимулирует ацинозные клетки, секретирующие ферменты.
Каждый из этих гормонов оказывает также слабое действие на другую систему, а
именно секретин действует на ацинусы, а ХЦК–на эпителий протоков. Вазоактивный
интестинальный полипептид (ВИП) по строению близок к секретину, а гастрин–к
ХЦК. Оба они обладают значительно более слабым действием, чем основные гормоны,
и конкурируют с ними за рецепторы: секретин–с ВИП, а гастрин–с ХЦК, что
приводит к взаимному конкурентному ингибированию. Слабой стимулирующей
активностью обладают также вещество Р и нейротензин. Панкреатические
полипептиды соматостатин и глюкагон угнетают секрецию панкреатического сока.
Нервная стимуляция осуществляется блуждающим нервом,
причем в качестве нейромедиатора наряду с ацетилхолином был идентифицирован
ВИП. Нервные стимулы вызывают, подобно ХЦК, выделение секрета, богатого
ферментами, и эта секреция может быть подавлена атропином.
Фазы панкреатической секреции. Базальная секреция бикарбоната и ферментов
составляет 2–3 и 10–15% максимального уровня соответственно. В цефалической
фазе, вызванной мыслями о еде, ее запахом, вкусом и актом глотания,
секреция бикарбоната повышается до 10–15%, а ферментов–до 25% максимального
уровня. Эта фаза связана с рефлекторным возбуждением блуждающего нерва, поэтому
может быть подавлена атропином или путем ваготомии. При поступлении пищи в
желудок начинается желудочная фаза, во время которой секреция
панкреатического сока еще более повышается под действием тех же стимулов
(активности блуждающего нерва и гастрина), которые вызывают
секрецию желудочного сока. Наиболее важная кишечная фаза панкреатической
секреции начинается с поступлением химуса в двенадцатиперстную кишку. При этом
S–клетки слизистой тонкого кишечника выделяют секретин, а I–клетки – ХЦК.
Адекватным стимулом для выделения секретина служит снижение рН ниже 4,5,
вызванное поступлением кислого содержимого желудка в проксимальный отдел
двенадцатиперстной кишки.
Соляная кислота потенциально может повреждать
слизистую тонкого кишечника, но такое воздействие предотвращается за счет двух
процессов–быстрой нейтрализации кислоты бикарбонатом, секретируемым
поджелудочной железой и слизистой двенадцатиперстной кишки под действием
секретина, а также, в меньшей степени, холецистокинина и абсорбции ионов Н+
Благодаря этим процессам рН содержимого двенадцатиперстной кишки повышается до
6–8 (диапазон рН, необходимый для действия панкреатических ферментов). Ионы H+,
секретируемые стимулированным желудком, должны при этом удаляться со скоростью
20–40 ммоль/ч.
Секрецию ХЦК эндокринными клетками слизистой тонкого кишечника
стимулируют продукты переваривания белков и жиров – пептиды, аминокислоты и
жирные кислоты, особенно длинноцепочечные. Углеводы таким действием не
обладают. Эта гормональная стимуляция дополняется рефлекторной стимуляцией
через блуждающий нерв.
Даже при полном наборе стимулов количество
секретируемого панкреатического сока составляет только 70% того максимального
количества, которое выделяется при внутривенном введении гормонов. Причиной
может служит выделение ингибирующих гормонов–соматостатина, панкреатического
полипептида и глюкагона.
Поджелудочная железа обладает большим запасом
функциональной активности. Она образует в 10 раз больше ферментов, чем
требуется для адекватного переваривания пищи, поэтому после удаления 90% железы
активности оставшихся 10% ткани достаточно для предотвращения несварения пищи.
Патофизиологические
аспекты. При нарушении функции
поджелудочной железы клинические симптомы появляются только на относительно
поздней стадии болезни. Признаками нарушения являются потеря в весе и жирный
стул, а причиной может быть, например, хронический панкреатит (как
правило, вследствие хронического алкоголизма) или карцинома поджелудочной
железы на поздней стадии. Недостаточность ферментов может быть частично
компенсирована пероральным приемом панкреатических ферментов.
Экзокринную функцию поджелудочной железы можно
оценить с помощью довольно трудоемкой пробы на секретин–ХЦК, при которой
вызывают максимальную стимуляцию секреции панкреатического сока и собирают его
через катетер. Более простым способом служит пероральное введение вещества
(например, флуоресцеина в панкреолауриновой пробе или парааминобензойной
кислоты в ПАБК–пробе), гидролизуемого панкреатическими ферментами до
конечных продуктов, которые всасываются и выделяются с мочой; по концентрации
этих продуктов в моче и оценивают эффективность панкреатической секреции.
29.5. Печень и желчная система
Печень–это самый крупный
орган тела, занимающий центральное место в обмене веществ. Она выполняет множество
функций, участвуя в обмене белков, углеводов, жиров, гормонов и витаминов, а
также в обезвреживании многих эндогенных и экзогенных веществ. Эти процессы
описаны в учебниках физиологической химии, а в настоящем разделе в связи с
пищеварительной системой мы рассмотрим лишь выделительную функцию печени,
т.е. секрецию желчи. Желчь состоит из воды, минеральных солей, слизи,
липидов холестерола и лецитина и двух видов специфических компонентов – желчных
кислот и пигмента билирубина. Желчные кислоты являются детергентами, и их
эмульгирующее действие играет важную роль в переваривании липидов.
Билирубин–это конечный продукт распада гемоглобина, подлежащий выведению из
организма.
Образование желчи
Функциональная анатомия. Клетки печени (гепатоциты)
образуют пластинки толщиной в одну клетку, разделенные узкими щелями (пространство
Диссе), представляющими собой заполненные
кровью синусоиды, эквивалентные капиллярам. В
стенках синусоидов имеются поры, через которые могут проходить такие крупные
макромолекулы, как альбумин и липопротеины. По мельчайшим канальцам –
желчным капиллярам, ограниченным плазматическими мембранами двух соседних
гепатоцитов,–желчь собирается в более крупные канальцы Геринга, стенки
которых, как и более крупных по размеру междольковых канальцев и желчных
протоков, образованы кубическими секреторными клетками. Мелкие канальцы
внутри долек печени и между ними сливаются в более крупные, образуя в конечном
счете печеночный проток. От этого протока отходит пузырный проток
к желчному пузырю. После слияния печеночный и пузырный протоки образуют общий
желчный проток, открывающийся в двенадцатиперстную кишку на вершине
фатерова сосочка позади протока поджелудочной железы или рядом с ним (рис.
29.1).
Функции
желчи. Желчь выполняет множество
важных функций. Вместе с ней выводятся конечные продукты обмена,
например билирубин, а также лекарственные препараты и токсины. Выделение с
желчью холестерола играет важную роль в регуляции его баланса. Желчные
кислоты необходимы для эмульгирования и всасывания жиров. Кроме того, желчь
содержит воду, минеральные соли и слизь. В сутки выделяется около 600 мл
желчи, и 2/3 этого количества поступает из канальцев, а 1/3–из более крупных
протоков.
Канальцевая желчь образуется приблизительно в
равных количествах при участии двух разных механизмов – зависимого от желчных
кислот и независимого от них (рис. 29.27).
Секреция, зависимая от желчных кислот. Установлена тесная
взаимосвязь между скоростью выделения желчи и секрецией желчных
кислот. В канальцевой желчи концентрация желчных кислот в 100 раз выше, чем
в портальной крови, поэтому считают, что они выделяются путем активного
транспорта с участием переносчика. Вслед за желчными кислотами по
осмотическому градиенту в канальцы устремляется вода, поэтому желчь изотонична
крови.
Существует два источника желчных кислот. Во–первых,
они синтезируются в самих гепатоцитах de novo из
холестерола при участии 7–гидроксилазы. Данный фермент играет ключевую роль и
по механизму обратной связи контролирует скорость синтеза желчных кислот.
Во–вторых, гепатоциты способны активно поглощать желчные кислоты из портальной
крови и выделять их в канальцы (см. также рис. 29.29). Эта экстракция очень
эффективна; при однократном прохождении крови через печень из нее извлекается
80% желчных кислот. Поэтому
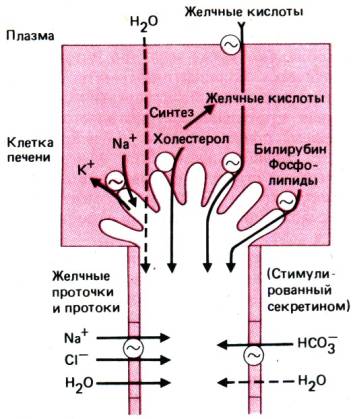
Рис. 29.27. Механизмы секреции желчи.
Секреция, зависимая от желчных кислот,– вверху справа. Желчные кислоты,
поглощаемые клетками печени из портальной крови или синтезированные в них
заново, активно транспортируются в канальцы. Секреция, независимая от желчных
кислот, вверху слева. Электролиты секретируются за счет активной
секреции ионов Nа+. Модификация первичной желчи (внизу):
ионы K+, Сl– и НСО3–
активно секретируются в желчные протоки, а вслед за ними под действием
осмотического давления выходит вода
концентрация желчных кислот
в периферической крови намного ниже, чем в воротной системе. Поскольку желчные
кислоты извлекаются из крови в 6 раз быстрее, чем поступают в канальцы, именно
последний процесс лимитирует скорость секреции желчных кислот.
Секреция, независимая от желчных кислот. В этом процессе участвуют
ионы Na+ , Cl–, НСО3–
и вода. Движущей силой служит активный транспорт Na+, возможно вместе с
бикарбонатом. Секрецию, независимую от желчных кислот, стимулирует, в частности,
секретин.
Помимо желчных кислот в канальцы активно
секретируются билирубин, холестерол и фосфолипиды (преимущественно лецитин)
(рис. 29.27). Нерастворимый в воде («непрямой») билирубин, большая часть
которого образуется из гемоглобина состарившихся эритроцитов, поступает в
гепатоциты в виде коллоидного агрегата, связанного с альбумином. Суточное
образование его составляет около 4 г/кг массы тела, или 200–300 мг/сут. В
гепатоцитах 80% билирубина конъюгировано с глюкуроновой кислотой и небольшое его
количество с серной кислотой. В такой конъюгированной форме билирубин
выделяется с желчью («прямой» билирубин). В основном тем же путем
выводятся лекарственные препараты и токсины.
Модификация желчи в желчных протоках
(рис. 29.27). В протоках, куда открываются канальцы,
первичная желчь подвергается модификации. Этот процесс напоминает процесс
модификации клубочкового фильтрата в почечных канальцах, и сходным образом для
него рассчитывается клиренс, только в случае желчи в качестве инертного
вещества используют вместо инулина эритритол или маннитол,
которые секретируются в канальцы, но не реабсорбируются. Подобные исследования
показали, что около 180 мл желчи, или 1/3 ее общего количества, выделяется в
протоки при активной секреции НСО3–. Этот процесс стимулируется
секретином.
Печеночная и пузырная желчь
Состав печеночной желчи (табл. 29.3). Желчь, выделяемая печенью со скоростью
0,4 мл/мин, имеет золотистый цвет, что объясняется присутствием в ней билирубина.
Концентрация электролитов в этой желчи такая же, как в плазме, за исключением
того, что она содержит в два раза больше НСО3– и
несколько меньше С1–. В то же время по составу органических веществ
желчь сильно отличается от плазмы, так как в желчи они представлены почти
исключительно желчными кислотами, холестеролом и фосфолипидами.
Таблица
29.3. Состав печеночной и пузырной желчи
|
Компоненты
|
Печеночная желчь,
ммоль/л
|
Пузырная желчь,
ммоль/л
|
|
|
|
Na+
|
165
|
280
|
|
K+
|
5
|
10
|
|
Са2+
|
2,5
|
12
|
|
С1–
|
90
|
15
|
|
НСО3–
|
45
|
8
|
|
Желчные кислоты
|
35
|
310
|
|
Лецитин
|
1
|
8
|
|
Желчные пигменты
|
0,8
|
3,2
|
|
Холестерол
|
3
|
25
|
|
рН
|
8,2
|
6,5
|
Желчные кислоты образуются в печени из холестерола в результате
его гидроксилирования и присоединения карбоксильной группы. Образующиеся в
печени кислоты – это первичные желчные кислоты;
к ним относятся хенодезоксихолевая
(диоксиформа) и холевая (триоксиформа) кислоты. В печени они
находятся не в свободной форме, а в виде конъюгатов с глицином и таурином,
причем конъюгатов с глицином образуется в три раза больше, поскольку количество
таурина ограничено. Конъюгированные желчные кислоты лучше растворимы в воде,
чем неконъюгированные, и обладают большей способностью к диссоциации и
образованию желчных солей с катионами, главным образом с ионами Na+ В кислой среде (рН 4,0)
соли желчных кислот нерастворимы и выпадают в осадок, но при физиологических
значениях рН (в тонком кишечнике) они хорошо растворимы.
В дистальном отделе подвздошной кишки и в толстой
кишке часть солей первичных желчных кислот подвергается дегидроксилированию под
действием анаэробных бактерий и превращается во вторичные желчные
кислоты–литохолевую (монооксиформа) и дезоксихолевую (диоксиформа).
Хенодезоксихолевая, холевая и дезоксихолевая кислоты присутствуют в соотношении
2:2:1. Литохолевая кислота присутствует лишь в некоторых фракциях, поскольку
большая ее часть экскретируется.
Эмульгирующее действие желчных кислот на жиры
основано главным образом на их способности образовывать мицеллы.
Молекулы желчных кислот имеют такую трехмерную структуру, при которой
гидрофильные карбоксильные и гидроксильные группы находятся на одной стороне
молекулы, а гидрофобная часть молекулы (стероидное ядро, метальные группы) на
противоположной, за счет чего молекулы желчных кислот обладают и гидрофильными,
и липофильными свойствами. Благодаря такому строению молекулы желчных
кислот действуют как детергенты: на границе раздела липидной и водной фаз они
образуют почти мономолекулярную пленку, в которой гидрофильные группы обращены
к водной, а липофильные – к липидной фазе. В водной фазе желчные кислоты
образуют упорядоченные агрегаты –мицеллы при условии, что их
концентрация достигает определенного уровня, называемого критической
концентрацией мицеллообразования (1–2 ммоль/л). Внутренняя, липофильная
область мицеллы может содержать липиды, например холестерол и
фосфолипиды; такие мицеллы называют смешанными (рис. 29.28). Сам по себе
холестерол нерастворим в воде, но в составе мицелл может находиться в растворе.
Если его концентрация превышает емкость мицелл, он образует кристаллический
осадок; этот процесс лежит в основе образования холестероловых желчных камней.
Состав пузырной желчи (табл. 29.3). Емкость
желчного пузыря составляет всего 50–60 мл. В то же время печень секретирует
желчь со скоростью 600 мл/сут, и половина этого количества перед поступлением в
тонкий кишечник проходит через желчный пузырь. Разница между объемом желчи,
поступающей в желчный пузырь, и его емкостью компенсируется высокоэффективной
реабсорбцией воды в желчном пузыре. В течение нескольких часов из желчи
может реабсорбироваться 90% воды. При
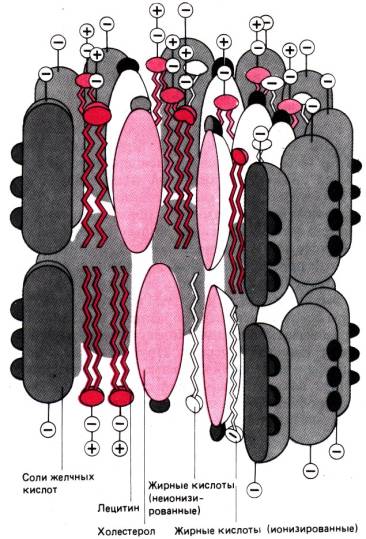
Рис. 29.28. Строение смешанной мицеллы. Сердцевина
мицеллы, состоящая из холестерола, лецитина, жирных кислот и моноглицеридов,
покрыта снаружи желчными кислотами, гидрофильные группы которых находятся на
поверхности мицеллы
этом органические вещества
остаются в желчном пузыре и их концентрация в желчи повышается. Движущей силой
реабсорбции служит активный транспорт ионов Nа+ при участии
«насоса», встроенного в базальную и латеральную мембраны клеток и активируемого
(Na+K+–ATФазой. Вслед за ионами Na+ перемещаются ионы С1–
и НСО3–, диффундирующие в направлении электрического
градиента или транспортируемые переносчиками. В результате реабсорбции НСО3–
рН пузырной желчи уменьшается до 6,5 против 8,2 в печеночной желчи. Вследствие
создания в межклеточном пространстве эпителия желчного пузыря высокой концентрации
ионов Na+ возникает осмотический градиент, приводящий к
накачиванию воды, которая оттекает затем в капилляры.
Моторика желчного пузыря. В состоянии натощак желчь скапливается в желчном
пузыре, а во время приема пищи выделяется в результате сокращений желчного
пузыря. Основным стимулятором сократительной активности желчного пузыря служит
холецистокинин, секретируемый слизистой оболочкой двенадцатиперстной кишки
при поступлении в нее химуса, содержащего жиры. До некоторой степени сокращения
желчного пузыря стимулируются также блуждающим нервом и парасимпатолитиками.
Они начинаются уже через 2 мин после того, как жирная пища соприкасается со
слизистой кишечника, и через 15–90 мин пузырь полностью опорожняется. Моторика
желчного пузыря включает два процесса. Вначале развивается тоническое
сокращение, вследствие чего уменьшается диаметр желчного пузыря, а затем на
этот эффект накладываются периодические сокращения, частота которых составляет
2–6/мин. В результате этих двух процессов создается давление 25–30 мм рт. ст.
Кишечно–печеночная циркуляция
Циркуляция желчных кислот (рис. 29.29). Желчные кислоты выделяются в
двенадцатиперстную кишку в виде смешанных мицелл. Несмотря на разведение
желчных кислот содержимым желудка, их концентрация в кишечнике составляет около
10 ммоль/л и остается выше критической концентрации мицеллообразования. Здесь в
добавление к холестеролу и лецитину в мицеллы включаются продукты
гидролитического расщепления жиров –жирные кислоты и моноглицериды.
При первоначальном контакте мицелл с кишечной стенкой
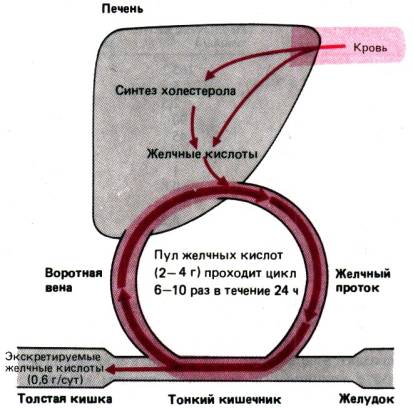
Рис. 29.29. Кишечно–печеночная
циркуляция желчных кислот
липиды диффундируют через мембрану щеточной каемки в
энтероциты, а желчные кислоты остаются в просвете кишечника, но при дальнейшем
прохождении по кишечнику желчные кислоты всасываются путем активного и
пассивного транспорта.
Около 50% желчных кислот
всасывается в кишечнике пассивным путем. В результате расщепления конъюгатов
желчных кислот и дегидроксилирования последних под действием кишечных бактерий
повышается их растворимость в липидах и облегчается пассивная диффузия.
Активное всасывание желчных кислот происходит
исключительно в концевом отделе подвздошной кишки–редкое явление, известное еще
только для всасывания витамина В12. Активному всасыванию подвергаются лишь те
желчные кислоты, которые обладают большой полярностью, затрудняющей их
пассивное всасывание, например конъюгаты таурина. Для процесса всасывания
желчных кислот в концевом отделе подвздошной кишки характерны типичные признаки
активного транспорта: кинетика насыщения и конкурентное ингибирование.
Небольшое количество желчных кислот (7–20%) не включается ни в активное, ни в
пассивное всасывание и выводится из организма.
Присутствие желчных кислот в
толстой кишке играет, по–видимому, важную роль в регуляции консистенции каловых
масс. При концентрации диоксижелчных кислот в толстой кишке свыше 3 ммоль/л в
просвет кишечника секретируется значительное количество электролитов и воды,
что приводит к поносу. Резко выраженная форма этого «хологенного» поноса
может наблюдаться при резекции или заболевании концевого отдела подвздошной
кишки, и для его лечения используют связывание желчных кислот с помощью
ионообменника холестирамина.
При поступлении поглощенных
желчных кислот в печень из них вновь образуются конъюгаты, а некоторые
вторичные желчные кислоты подвергаются гидроксилированию. Потеря желчных кислот
с калом (0,2–0,6 г/день) компенсируется за счет их синтеза.
Общий пул желчных кислот в организме составляет
около 3,0 г. Этого количества недостаточно для обеспечения липолиза после
приема пищи; в частности, при потреблении жирной пищи желчных кислот требуется
в 5 раз больше. Однако организм не испытывает дефицита в желчных кислотах,
поскольку они многократно циркулируют через кишечник и печень (кишечно–печеночная
циркуляция). Частота, с которой пул желчных кислот совершает полный цикл,
зависит от пищевого режима и составляет от 4 до 12 циклов в сутки.
Циркуляция билирубина. Желчный пигмент билирубин, подобно желчным
кислотам и липидам, поступает в кишечник в форме глюкуронида. Лишь небольшое
количество этого полярного соединения реабсорбируется в желчном пузыре и тонком
кишечнике. В концевом отделе подвздошной кишки и (в основном) в толстой кишке
конъюгат билирубина расщепляется под действием бактериальных гидролаз. При этом
билирубин превращается в уробилиноген, который наряду с другими
продуктами распада билирубина придает коричневый цвет каловым массам. Менее 20%
уробилиногена всасывается обратно, и из этого количества около 90% вновь
попадает в печень и возвращается в желчь, а остальные 10% экскретируются с
мочой.
Патофизиологические аспекты. Повышенное содержание
уробилиногена в моче может указывать на заболевание печени, сопровождающееся
нарушением экскреции билирубина. Полное отсутствие уробилиногена в моче,
светлый цвет кала и желтушность указывают на полную закупорку желчного
пузыря; в этой ситуации билирубин совсем не поступает в кишечник и
уробилиноген не образуется.
Наиболее известное и
распространенное нарушение нормальной физиологии желчной системы – это
выпадение в осадок холестерола с образованием холестероловых желчных камней.
Холестерол, как и лецитин, находится в растворенном состоянии только в составе
смешанных мицелл. Если возрастает концентрация холестерола либо становится
ниже критического уровня концентрация желчных кислот или лецитина,
холестерол выпадает в осадок. К числу факторов, вызывающих повышение
относительного содержания холестерола. относятся эстрогены, углеводная диета,
избыточная масса и процессы, снижающие концентрацию желчных кислот, например
воспаление подвздошной кишки (болезнь Крона) или ее резекция. В
некоторых случаях достаточно бывает перорального приема желчных кислот, чтобы
литогенная желчь превратилась в алитогенную, в которой холестероловые камни
могут раствориться. Больше всего для этой цели подходят хенодезоксихолевая и
уродезоксихолевая кислоты, так как они не вызывают поноса.
Клиническим проявлением
нарушенного обмена билирубина является желтуха. Желтый цвет кожи связан
с повышенным содержанием в плазме билирубина, которое может иметь место в
следующих случаях:
1) при повышенном образовании билирубина в результате
усиленного распада эритроцитов (гемолитическая желтуха);
2) в результате нарушения процесса конъюгирования или
транспорта билирубина в гепатоцитах, как, например, при желтухе беременных
или родильной желтухе Джильберта;
3) при задержке оттока желчи, например из–за желчных
камней или опухолей, локализованных в области желчного протока (механическая
желтуха).
29.6. Тонкий кишечник
Тонкий кишечник выполняет
несколько важных функций:
1) перемешивание химуса с секретами
поджелудочной железы, печени (желчью) и слизистой кишечника;
2) переваривание пищи;
3) всасывание гомогенизированного и
переваренного материала;
4) дальнейшее продвижение оставшегося
материала по желудочно–кишечному тракту;
5) секреция гормонов;
6) иммунологическая
защита.
Тонкий кишечник включает три
отдела–двенадцатиперстную кишку (длиной 20–30 см), тощую кишку,
начинающуюся от перетяжки Трейтца и имеющую в длину 1,5–2,5 м, и подвздошную
кишку (длиной 2–3 м), в которую тощая кишка переходит без четкой границы.
Общая длина тонкого кишечника составляет около 4 м в состоянии тонического
напряжения (при жизни) и около 6–8 м в атоническом состоянии (после смерти).
Двигательная активность
тонкого кишечника состоит из непропульсивных перемешивающих движений и пропульсивчой
перистальтики. Она зависит от собственной активности гладкомышечных клеток,
а также от влияния вегетативной нервной системы и многочисленных гормонов, в
основном желудочно–кишечного происхождения.
Основной миогенный ритм. Сокращения мышц тонкого кишечника, так же как и
желудка, определяются основным миогенным ритмом; на эти медленные
волны накладывается потенциал действия. В верхнем отделе кишечника
пейсмекеры медленных волн обладают большей частотой (12 циклов в минуту), чем в
подвздошной кишке, по длине которой частота сокращений постепенно уменьшается
до 8 циклов в минуту. Благодаря наличию этого орально–анального градиента
содержимое кишечника медленно продвигается по кишечнику даже во время
непропульсивной перистальтики. Кроме того, в верхнем отделе тонкого кишечника
возбудимые мышечные клетки расположены более плотно, чем в дистальных отделах,
и возникающий сдвиг фаз оказывает такое же воздействие, как и градиент частоты
сокращений (рис. 29.19).
Нервная и гормональная регуляция. Как во внутренней, так и во
внешней нервной регуляции моторики тонкого кишечника особенно важную роль
играет ауэрбахово сплетение. Медиатором в этой системе служит
ацетилхолин, угнетающий активность слоя циркулярных мышц, от которой
зависит двигательная активность тонкого кишечника. Если устранить это
угнетающее действие (например, в эксперименте путем блокады потенциалов
действия нервных волокон тетродотоксином, не оказывающим иного воздействия на
гладкие мышцы), кишечник начинает сильно сокращаться с частотой медленных волн.
В физиологических условиях временное прекращение тормозного действия
ацетилхолина, по–видимому, обеспечивается вазоактивным интестинальным
полипептидом. На слой продольных мышц ацетилхолин оказывает противоположное
действие, т.е. стимулирует его сокращения. Внешняя иннервация играет в
регуляции моторики тонкого кишечника второстепенную роль. Симпатические
нервные волокна, выходящие из сегментов Т9–10 спинного мозга и из синапсов
чревного и брыжеечного ганглиев, угнетают двигательную активность тонкого
кишечника, а парасимпатическая система (блуждающий нерв) стимулирует ее.
В регуляции моторики тонкого кишечника участвуют
также многие гормоны, которые могут обладать паракринным, эндокринным
или нейрокринным действием и оказывать стимулирующее или угнетающее влияние.
Механизмы регуляции очень сложны, и до сих пор ни один из видов двигательной
активности кишечника не удалось связать с действием какого–либо определенного
гормона. Нервные и гормональные эффекты индуцируются приемом пищи и растяжением
кишечника.
Последовательность движений тонкого кишечника. Движения кишечника в состоянии
натощак отличаются от таковых в пищеварительной фазе. В первом
случае преобладает направленный пропульсивный характер миоэлектрического
двигательного комплекса.
Прием пищи прерывает двигательную активность, способствующую
продвижению химуса. В пищеварительной фазе преобладают ритмическая
сегментация и маятникообразные движения, способствующие перемешиванию
содержимого кишечника. Это изменение характера двигательной активности вызывают
желудочно–кишечные гормоны гастрин и холецистокинин.
Продолжительность и интенсивность двигательной активности в пищеварительной
фазе зависят от состава пищи и ее калорийности. Кроме того, при одинаковом
энергетическом потенциале триглицериды действуют значительно сильнее и в
течение более продолжительного времени по сравнению с углеводами и белками.
В результате редких периодических пропульсивных
движений и сдвига фаз медленных волн содержимое кишечника медленно
передвигается в направлении толстого кишечника. При средней скорости
перемещения 1–4 см/мин пища достигает слепой кишки за 2–4 ч. На продолжительность
эвакуации пищи влияет ее состав. В зависимости от состава пищи скорость ее
перемещения уменьшается в ряду: углеводы, белки, жиры. Кроме того, в кишечнике
существует двигательная активность меньшего масштаба –сокращения ворсинок,
способствующие перемешиванию пищи и взбалтыванию неперемешивающегося слоя.
Ворсинки ритмично
укорачиваются в такт сокращениям мышечной
пластинки, и частота их сокращений также уменьшается в направлении от
проксимальных отделов кишечника к дистальным, а наибольшая активность
наблюдается в двенадцатиперстной кишке. Сокращения ворсинок способствуют также
опорожнению центрального лимфатического протока. Индуцирует их гормон вилликинин,
образующийся в слизистой оболочке тонкого кишечника.
Илеоцекальная заслонка. Тонкий кишечник заканчивается участком длиной около
4 см, который контролирует эвакуацию пищи в толстый кишечник. Обычно этот
илеоцекальный сфинктер находится в состоянии тонического сокращения, и
давление в нем составляет около 20 мм рт. ст. При растяжении концевого участка
подвздошной кишки сфинктер расслабляется, а с увеличением давления в слепой
кишке сокращается (рис. 29.30). Кроме того, в месте перехода подвздошной кишки
в слепую имеются две полулунные складки, образующие илеоцекальный клапан,
который выдерживает давление до 40 мм рт. ст. Благодаря такому анатомическому
барьеру численность бактерий в подвздошной кишке в 105 раз
меньше, чем в слепой (см. рис. 29.40). На этой разнице основан метод
определения времени транзита пищи через тонкий кишечник и перехода ее в
слепую кишку. Применяемый в этом методе невсасывающийся углевод, пройдя по
тонкому кишечнику и поступив в слепую кишку, разлагается под действием бактерий
с образованием довольно большого количества Н2, который определяют в
выдыхаемом воздухе.
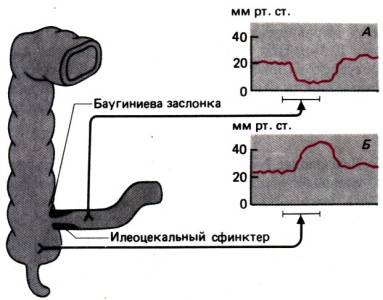
Рис. 29.30. Зависимость функции
илеоцекального сфинктера (баугиниевой заслонки) от давления. При
повышении давления в подвздошной кишке тонус илеоцекальной заслонки
уменьшается, а при повышении давления в слепой кишке–увеличивается
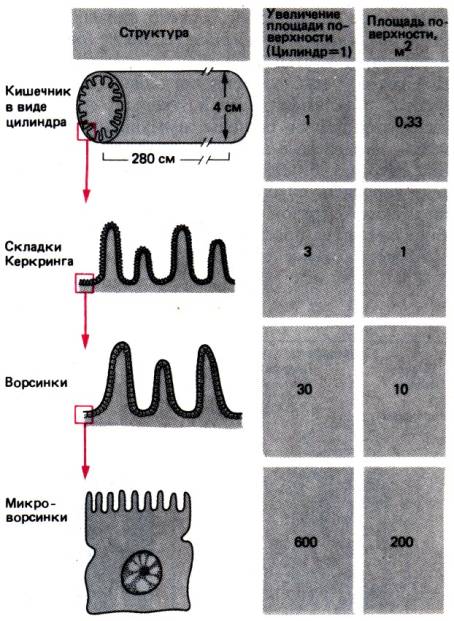
Рис. 29.31. Увеличение поверхности
слизистой оболочки за счет особенностей морфологии
Всасывающая поверхность и
кровоток.
Наличие складок и ворсинок обеспечивает большую всасывающую поверхность тонкого
кишечника. Как показано на рис. 29.31, за счет круговых складок, называемых складками
Керкринга, ворсинок и микроворсинок, всасывающая поверхность
цилиндрической трубки увеличивается в 600 раз и достигает 200 м2.
Функциональную единицу образуют ворсинка с ее внутренним содержимым и
лежащими под ней структурами и крипта, разделяющая соседние ворсинки
(рис. 29.32). Эпителий тонкого кишечника относится к тканям с наиболее высокой
скоростью деления и обновления клеток. Недифференцированные цилиндрические
клетки образуются в глубине крипты и мигрируют затем к вершине ворсинки; это
перемещение занимает 24–36 ч. По пути клетки созревают, синтезируют
специфические ферменты и транспортные системы (переносчики), необходимые для
всасывания и, достигая вершины ворсинки, представляют собой полностью
сформированные энтероциты. Всасывание компонентов пищи происходит
главным образом в верхней части ворсинки, а секреторные процессы–в криптах.
Помимо энтероцитов в слизистой тонкого кишечника присутствуют слизистые
клетки, а также различные эндокринные клетки, называемые аргентаффинными
в связи с тем, что они поглощают кристаллы серебра. С лимфатической тканью
желудочно–кишечного тракта связаны иммунокомпетентные клетки, называемые в
связи с их формой М–клетками. Через 3–6 дней клетки, находящиеся на
вершине ворсинки, слущиваются и заменяются новыми. Таким образом в течение
нескольких дней обновляется вся поверхность кишечника.Кровоснабжение
слизистой тонкого кишечника
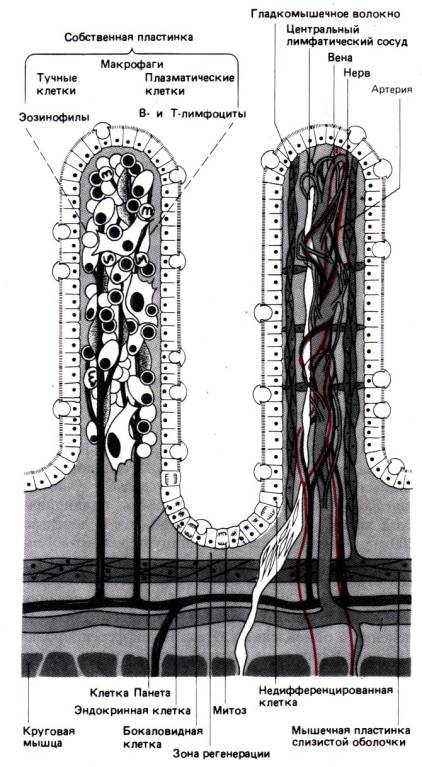
Рис. 29.32. Поперечное сечение двух
ворсинок тонкого кишечника и крипты между ними, на котором видны несколько
типов клеток слизистой и структуры, находящиеся внутри ворсинки
обеспечивает
в основном верхняя брыжеечная артерия, но двенадцатиперстная кишка снабжается
чревной артерией, а концевой отдел подвздошной кишки–нижней брыжеечной
артерией. Ответвления этих сосудов образуют центральные сосуды ворсинок
(рис. 29.32), которые разветвляются на субэпителиальные капилляры. На тонкий
кишечник приходится 10–15% крови, составляющей ударный объем сердца. Примерно
75% этого количества поступает в слизистую оболочку, около 5%–в подслизистую и
20%–в мышечный слой слизистой. После приема пищи кровоток увеличивается на
30–130% в зависимости от характера и объема пищи. Он распределяется таким
образом, что повышенный приток крови всегда направлен к участку, где в данный
момент находится основная масса химуса.
Всасывание
воды. В среднем за сутки через
тонкий кишечник проходит около 9 л жидкости. Приблизительно 2 л поступают
из крови и 7 л–с эндогенными секретами желез и слизистой кишечника (рис.
29.33). Более 80% этой жидкости всасывается обратно в тонком кишечнике–около
60% в двенадцатиперстной кишке и 20% в подвздошной кишке. Остальная жидкость
всасывается в толстом кишечнике и только 1%, или 100 мл, выделяется из
кишечника с каловыми массами.
Движение воды через слизистую всегда сопряжено с
переносом растворенных в ней веществ–несущих и не несущих заряда. Слизистая
верхних отделов тонкого кишечника относительно проницаема для растворенных
веществ. Эффективный размер пор в этих отделах составляет около 0,8 нм (ср. 0,4
нм в подвздошной кишке и 0,23 нм в толстой кишке), поэтому в том случае, когда
осмолярность химуса в двенадцатиперстной кишке отличается от осмолярности крови,
данный параметр выравнивается в течение нескольких минут (рис. 29.34). При
гиперосмолярности химуса вода поступает в просвет кишечника, а при его
гипоосмолярности быстро всасывается. В процессе дальнейшего прохождения по
кишечнику химус остается изотоничным плазме.
Всасывание Na+ (рис.
29.35). Одна из чрезвычайно важных функций тонкого кишечника – это транспорт
ионов Na+ Именно за счет ионов Na+ создаются
в основном электрический и осмотический градиенты; кроме того, ионы Na+ участвуют в сопряженном транспорте
других веществ. Всасывание Na+ в кишечнике происходит очень
эффективно: из 200–300 ммоль Na+ , ежедневно поступающих в
кишечник с пищей, и 200 ммоль секретируемого в него Na+ с калом выводятся только 3–7 ммоль, основная же часть Na+ всасывается в тонком
кишечнике.Всасывание ионов Na+ в кишечнике происходит
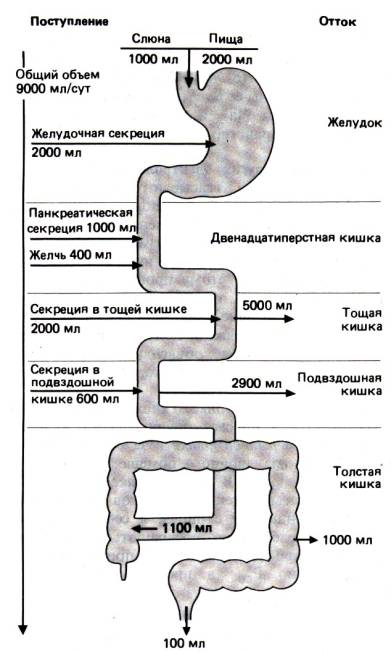
Рис. 29.33. Баланс жидкости в желудочно–кишечном тракте. Из
общего количества жидкости, поступающей в желудочно–кишечный тракт с пищей (2
л) и эндогенными секретами (7 л), с калом выводится только 100 мл
как за счет активного, так и
за счет пассивного механизмов, в том числе путем электрогенного транспорта,
транспорта, сопряженного с переносом незаряженных соединений (котранспорт,
например глюкозы, аминокислот), электронейтрального транспорта NaCI, (Na+–H+–обмена и конвекции
(следование за растворителем).
При электрогенном транспорте ионы Na+
переносятся через базолатеральную область мембраны в межклеточное пространство
с помощью натриевого насоса, получающего энергию за счет гидролиза АТФ
под действием (Na+K+–ATФазы (рис. 29.35/1). Это
главный механизм всасывания ионов Na+ в кишечнике. Перенос Na+ в данном случае происходит
против концентрационного градиента (концентрация Na+ в клетке составляет 15,
а в плазме–100 мМ) и против электрического
градиента (электрический заряд внутри клетки равен — 40 мВ, а в
межклеточном пространстве + 3 мВ).
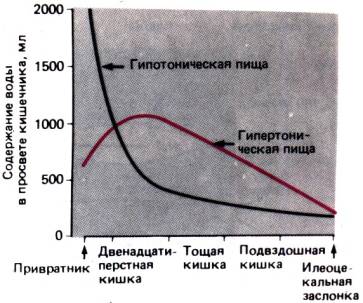
Рис. 29.34. Содержание воды в тонком кишечнике как функция
осмолярности пищи. После приема гипотонической пищи вода быстро всасывается из
просвета кишечника, а после приема гипертонической пищи вода выходит в просвет
кишечника для восстановления осмотического равновесия (по [22])
Отрицательный заряд внутри
клетки обусловлен тем, что на каждые три иона Na+ выводимых из клетки, в нее
поступают только два иона К+. Наличие этих двух градиентов
способствует поступлению Na+ в клетку из просвета кишечника.
Активность (Na+K+–ATФазы, а следовательно, и
активный транспорт Na+ могут быть подавлены с помощью сердечного
гликозида оубаина. В верхнем отделе тонкого кишечника из–за довольно
значительной проницаемости плотных контактов часть поглощенных ионов Na+ может выходить обратно в
просвет кишечника, и, если концентрация Na+ в просвете кишечника
составляет менее 133 мМ, всасывания фактически не происходит. Слизистая
подвздошной кишки является более «плотной», поэтому поглощение ионов Na+
в ней продолжается даже в том случае, если его концентрация в просвете
кишечника составляет 75 мМ.
Сходная ситуация имеет место
и при сопряженном транспорте ионов Na+ (рис. 29.35/2). В этом
случае незаряженные вещества (D–гексозы, L–аминокислоты, водорастворимые
витамины, а в подвздошной кишке и желчные кислоты) переносятся в клетку вместе
с ионами Na+ общими переносчиками. Таким образом,
активный транспорт Na+ через
базолатеральную область мембраны косвенным путем обеспечивает энергией процесс
всасывания органических веществ.
При электронейтральном
транспорте NaCI в клетку одновременно переносятся ионы Na+ и С1–, в результате чего процесс и является
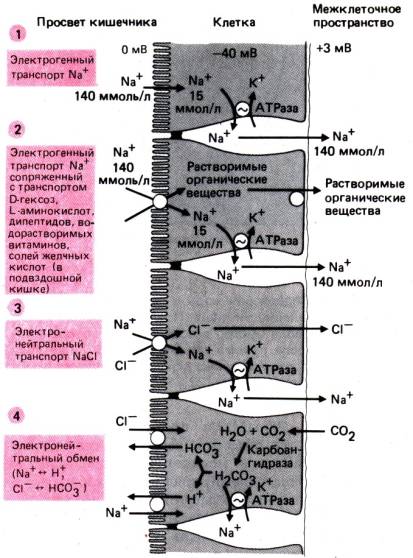
Рис. 29.35. Поглощение ионов в тонком
кишечнике. 1. Электрогенное поглощение ионов Na+ против
электрохимического градиента. 2. Сопряженный электрогенный транспорт Na+
(сопряженный с переносом органических веществ общим переносчиком). 3.
Нейтральный сопряженный транспорт. 4. Нейтральное поглощение Na+Cl– путем двойного обмена на
ионы Н+ и НСО3–, (особенно выражен в
подвздошной кишке). Источником энергии для всех четырех механизмов переноса
служит (Nа+К+–АТРаза (АТФаза) в
базальной и латеральной областях мембраны
электронейтральным (рис. 29.35/3). Повышение
концентрации ионов Са2+ или
цАМФ приводит к угнетению этого механизма, а если при этом происходит активная
секреция С1–, то в конечном итоге начинаются чистое выделение воды и
понос. Другое объяснение электронейтрального транспорта основано на
предположении о двойном обмене, при котором ионы Na+ обмениваются на ионы H+, а ионы С1––на
ионы НСО3 (рис. 29.35/4); при этом ионы Н+ и НСO3– образуются из Н2О
и СO2 .
Движущей силой и в этом случае служит активный транспорт ионов Na+
через базолатеральную область мембраны.
Исключительно важную роль во всасывании ионов Na+ в тонком кишечнике играет
пассивный транспорт путем конвекции. Благодаря довольно значительной
проницаемости эпителия до 85% ионов Na+ поглощается по механизму
«следования за растворителем». При определенной концентрации глюкозы ее
всасывание создает ток воды, с которым ионы Na+ и переносятся через
межклеточное пространство.
Всасывание других
электролитов. Ионы
К+ в отличие от ионов Na+ всасываются преимущественно за
счет пассивного транспорта по градиенту концентрации, поскольку концентрация
ионов К+ в клетке равна 14 мМ, а в плазме–4 мМ.
Ионы С1– всасываются частично вместе с ионами Na+
(см. выше); этому процессу способствует трансэпителиальный электрический
градиент, поскольку по отношению к просвету кишечника серозная поверхность
заряжена положительно. Существует интересная модель, объясняющая происхождение
некоторых видов диареи активной электрогенной секрецией ионов Cl–.
В верхнем отделе тонкого кишечника бикарбонат секретируется
в просвет бруннеровыми железами в двенадцатиперстной кишке и за счет
описанного выше механизма двойного обмена (рис. 29.35/4) в подвздошной
кишке. В тощей кишке ионы НСO3–, напротив, всасываются.
Часть ионов НСO3– , поступающих в кишечник с пищей и секретируемых в
верхнем отделе, может превращаться в СO2 под действием карбоангидразы. Этот процесс приводит к повышению
РСО2 в просвете кишечника до 300 мм рт. ст. и диффузии СО2
в клетки. Вследствие этого в верхнем отделе тонкого кишечника направление
двойного обмена противоположно тому, которое показано на рис. 29.35/4, – СO2 переносится из просвета кишечника в клетку, а ионы НСО3–
выходят в плазму, т.е. всасываются.
Ежедневно с молоком и молочными продуктами человек
потребляет около 1 г кальция. В кислой среде желудка соли кальция
диссоциируют и растворяются; около 40% ионов Ca2+ всасывается в тонком
кишечнике, главным образом в верхнем его отделе. При низких концентрациях Са2+
всасывание происходит путем активного транспорта, а при высоких
концентрациях включается и механизм пассивного транспорта. Как вход ионов Са2
+ в клетку, так и их выход из нее—это активные процессы;
в переносе участвует специфический Са2+–связывающий
белок в щеточной каёмке, и транспорт осуществляется через базолатеральную
область мембраны энтероцита при участии насоса, активируемого Са2+–АТФазой.
В регуляции всасывания ионов Са2+ участвуют паратгормон и (витамин
D)–гормон (1,25–дигидроксикальциферол).
Баланс железа в организме целиком зависит от его всасывания в
кишечнике, так как специального механизма регуляции его выведения не
существует. Из 10–20 мг железа, ежедневно поступающих с пищей, всасывается
только 10%. При дефиците железа его поглощение увеличивается в два и более раз.
В составе гемоглобина железо поглощается легче, чем в составе растительных
остатков, поскольку в последнем случае оно нередко присутствует в виде
нерастворимых соединений. Всасывание железа происходит в основном в верхнем отделе
тонкого кишечника, при этом оно поглощается как в двухвалентной, так и
(несколько медленнее) в трехвалентной форме. Поглощение и высвобождение
энтероцитами железа, как и Ca2+,–это процессы активного
транспорта. Выход Са2+ из энтероцитов происходит более медленно,
и именно данный процесс является лимитирующей стадией процесса поглощения
железа. В сыворотке крови железо связано с транспортным белком трансферрином и
в таком виде доставляется к месту его действия. Избыток железа связывается в
энтероцитах с ферритином и при слущивании состарившихся клеток поступает вместе
с ними в просвет кишечника и выводится.
Переваривание
и всасывание углеводов
В западных странах суточный
рацион человека включает 250–280 г углеводов; по калорийности это их количество
эквивалентно 4,3–13,7 кДж/г. Большая часть (около 60%) углеводов в пище
представлена растительным крахмалом – полисахаридом с мол. массой
100000–1000000. Около 30% углеводов пищи составляет сахароза, получаемая
из сахарного тростника или сахарной свеклы и используемая в виде сахара.
Примерно 10% углеводов пищи составляет лактоза. Помимо этих двух
дисахаридов в пище содержатся небольшие количества моносахаридов – глюкозы
и фруктозы, а также крахмал животного происхождения –гликоген.
Крахмал состоит из цепей
полимеризованной глюкозы. Амилоза (около 20% в составе крахмала)
представляет собой неразветвленные цепи, в которых первый углеродный атом
каждой молекулы глюкозы связан с 4–м углеродным атомом следующего остатка
глюкозы (a–1,4–гликозидная связь). В амилопектине
(приблизительно 80% в составе крахмала) через каждые 25 остатков глюкозы от
главной цепи отходит боковая цепь, так что 6–й углеродный атом молекулы глюкозы
главной цепи связан с первым углеродным атомом глюкозы в боковой цепи также a–1,4–гликозидной
связью. Гликоген имеет сходное строение, но в нем боковые цепи отходят
от каждого двенадцатого остатка глюкозы.
Ферментативный гидролиз. Присутствующая в составе слюны и панкреатического сока a–амилаза расщепляет a–1,4–гликозидные связи в
молекуле крахмала, но не действует на b–1,4–гликозидные связи,
например, в молекуле целлюлозы. Конечные продукты гидролиза под действием a–амилазы–мальтоза.
мальтотриоза, а в случае разветвленных амилопектинов–декстрины. Оптимум рН для a–амилазы–7,1.
Амилаза слюны расщепляет до
50% крахмала в составе пищи, если та достаточно долго пережевывается и
наслаивается затем на дне желудка таким образом, что фермент не может
инактивироваться под действием соляной кислоты. Чрезвычайно быстрый гидролиз
крахмала происходит в двенадцатиперстной кишке под действием больших количеств
амилазы, секретируемой поджелудочной железой. Небольшая часть панкреатической
амилазы действует, будучи связанной с поверхностью слизистой оболочки, но
физиологическое значение этого мембранного переваривания относительно невелико,
поскольку в просвете кишечника содержится в 10 раз больше амилазы.
Углеводы могут всасываться в кишечнике только в виде
моносахаридов, поэтому продукты, образовавшиеся под действием амилазы, должны
подвергаться дальнейшему гидролитическому расщеплению. Это расщепление
происходит в мембранах щеточной каемки под действием олигосахаридаз–группы
активных гидролитических ферментов, локализованных на поверхности, которая
обращена в просвет кишечника. Концентрация олигосахаридаз наиболее высока в
тощей кишке и значительно ниже–в двенадцатиперстной. a–1,6–связи в амилопектине и гликогене
расщепляются a–1,6–гликозидазой, также локализованной в
щеточной каемке. После приема больших количеств олигосахаридов в течение 2–5
дней происходит адаптация путем повышения концентрации ферментов. Активность
мембраносвязанных ферментов чрезвычайно высока, поэтому лимитирующим звеном в
усвоении углеводов является не их расщепление, а всасывание моносахаридов.
Исключение составляет лактоза, гидролиз которой происходит медленнее,
чем всасывание конечного продукта. Кроме того, довольно широко распространено
врожденное нарушение гидролитического расщепления лактозы, которое проявляется
в виде диареи, связанной с осмотическим действием невсосавшейся лактозы.
Всасывание моносахаридов. Конечными продуктами расщепления углеводов являются
глюкоза, галактоза и фруктоза (рис. 29.36). Альдогексозы глюкоза
и галактоза всасываются путем активного транспорта, сопряженного с переносом Na +, а всасывание фруктозы
носит пассивный характер и происходит путем облегченной диффузии. Глюкоза и
галактоза также могут всасываться путем пассивного переноса в случае очень
высокой их концентрации в просвете кишечника, какая обычно создается при
употреблении богатой углеводами пищи. Всасывание гексоз происходит относительно
быстро, хотя и медленнее, чем гидролитическое расщепление углеводов. В отличие
от крахмала
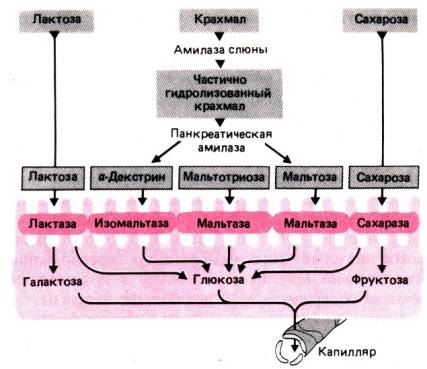
Рис. 29.36. Переваривание и всасывание
углеводов. Ферментативный гидролиз ди– и олигосахаридов в щеточной каемке.
Конечные продукты гидролитического расщепления углеводов панкреатическими
ферментами и дисахариды, поступающие с пищей, расщепляются до моносахаридов в
мембранах клеток щеточной каемки. Мономерными компонентами трех углеводов в
центре рисунка служат только молекулы глюкозы (по [12])
конечные продукты его
расщепления обладают осмотической активностью, но за счет их быстрого
всасывания не происходит образования гипертонического раствора. Последовательность
стадий гидролитического расщепления углеводов и всасывания продуктов
расщепления приведена на рис. 29.36.
Переваривание и всасывание белков
Взрослые потребляют с пищей
70–90 г белков в день, а детям в расчете на 1 кг веса их требуется в 5–10 раз
больше. Почти такое же количество белков поступает в просвет кишечника в
составе пищеварительных соков, слущивающихся клеток и в виде белков сыворотки.
При синдроме экссудативной энтеропатии через кишечник теряется так много
белка плазмы, что его убыль не компенсируется синтезом новых белков в печени, и
возникает гипопротеинемия.
Ферментативный
гидролиз.
Переваривание белков начинается в желудке, но роль этого этапа относительно
невелика, поскольку пепсином гидролизуется не более 10–15% белков пищи.
У больных, страдающих ахилией и дефицитом пепсина, белки тем не менее могут
нормально перевариваться, так как в тонком кишечнике переваривание белков
происходит чрезвычайно эффективно. Через 10–20 мин после приема пищи начинается
образование панкреатических пептидаз, которое продолжается до тех пор,
пока белки присутствуют в кишечнике. Часть этих ферментов выделяется с калом.
Содержание химотрипсина в каловых массах может служить показателем
функциональной активности поджелудочной железы.
Различные панкреатические
ферменты, катализирующие гидролиз белков, атакуют белковую молекулу в разных
участках (табл. 29.2). В двенадцатиперстной кишке, куда эти ферменты поступают
в неактивной форме, происходит их активация трипсином, образующимся из трипсиногена
под действием энтерокиназы. Ферменты, гидролизующие белки, подразделяют на
эндопептидазы (трипсин, химотрипсин, эластаза) и экзопептидазы
(карбопептидазы А и В). Эндопептидазы расщепляют внутренние связи в белковой
молекуле с образованием олигопептидов, а экзопептидазы отщепляют аминокислоты с
концов пептидной цепи. Около 30% конечных продуктов гидролиза составляют
нейтральные и основные аминокислоты и 70% – олигопептиды,
состоящие из 2–6 аминокислотных остатков.
В щеточной каемке и внутри
энтероцитов присутствуют другие пептидазы. В цитозоле подвергается гидролизу
примерно 90% олигопептидов (ди–и трипептиды), поступающих в клетку при участии
специальных транспортных систем. Около 10% олигопептидов, главным образом
состоящих из 4–8 аминокислотных остатков, гидролизуют ферменты, локализованные
в щеточной каемке. В каждом случае итог процесса – это появление в крови
воротной вены аминокислот как конечных продуктов гидролитического
расщепления белков.
Всасывание белков, пептидов и аминокислот. Примерно 50–60% белков пищи
всасывается в двенадцатиперстной кишке и около 30%–по мере прохождения химуса
до подвздошной кишки, т. е. 80–90% экзогенных и эндогенных белков всасывается в
тонком кишечнике. Только около 10% белков достигает толстого кишечника, где они
расщепляются под действием бактерий. Небольшое количество белка выделяется с
калом, но эта часть белка приходится на слущивающиеся клетки, а не на
непереваренные остатки пищи.
Интактные молекулы белка поглощаются в очень
небольшом количестве путем пиноцитоза. Всасывание по этому пути не имеет
значения для усвоения белков, но может играть важную роль в связи с
иммунореактивностью, приводя к сенсибилизации и аллергии. Пептиды
всасываются в виде ди– и трипептидов путем пассивного переноса или активного
транспорта с участием переносчиков. Поглощение аминокислот происходит с
помощью четырех основных групп транспортных систем: для нейтральных,
двухосновных и дикарбоновых аминокислот и для иминокислот.
К дополнительной группе относится система для глицина. Системы первых
трех групп осуществляют перенос по механизму сопряжения с транспортом Na+, описанному выше; при этом
всасывание кислых дикарбоновых аминокислот происходит как пассивный процесс,
хотя и с участием переносчиков. За счет внутриклеточного переаминирования с
участием аланина концентрация этих аминокислот в клетке сохраняется низкой.
Различные аминокислоты одной группы ингибируют перенос друг друга, конкурируя
за один и тот же переносчик (конкурентное ингибирование).
Переваривание и всасывание нуклеопротеинов. Нуклеопротеины гидролизуются
и всасываются так же, как другие белки. Нуклеиновые кислоты–ДНК и РНК –
гидролизуются специальными
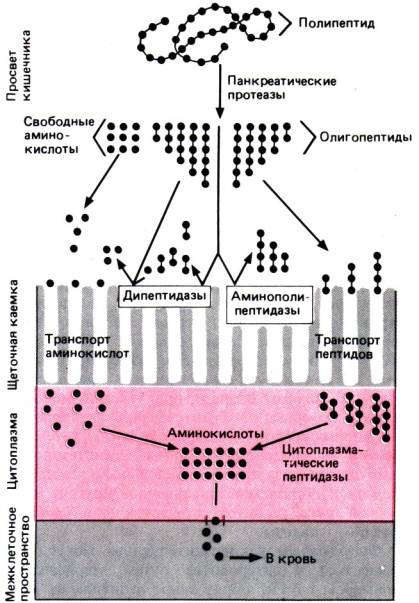
Рис. 29.37. Переваривание и всасывание
белков. Просвет кишечника: расщепление полипептидов до олигопептидов, ди– и
трипептидов и аминокислот. Мембраны клеток щеточной каемки: дальнейшее
расщепление специфическими пептидазами и поглощение аминокислот и
олигопептидов. Цитоплазма: расщепление ди– и олигопептидов цитоплазматическими
пептидазами до аминокислот. Базальная мембрана: выход аминокислот из клетки в
кровь
панкреатическими
ферментами – дезоксирибонуклеазой и рибонуклеазой и расщепляются
в щеточной каемке фосфодиэстеразами и нуклеотидазами до нуклеотидов.
Нуклеотиды транспортируются в энтероциты при участии специальных механизмов
(рис. 29.37).
Переваривание
и всасывание липидов
Человек потребляет в день
около 60–100 г жиров. Примерно 90% жиров пищи–это триглицериды, большую
часть которых составляют липиды, содержащие жирные кислоты с длинной цепью
– из 16 (пальмитиновая кислота) или 18 (стеариновая, олеиновая, линолевая
кислоты) атомов углерода. Триглицериды, содержащие жирные кислоты с короткой
цепью (2–4 углеродных атома) или средней цепью (6–8 атомов), составляют
лишь небольшую часть жиров пищи. Остальные 10% жиров пищи приходятся на
фосфолипиды (главным образом лецитин), эфиры холестерола и жирорастворимые
витамины (рис. 29.38).
Переваривание жиров. В желудке жиры образуют капельки диаметром около
100 нм. В щелочной среде тонкого кишечника при наличии белков, продуктов
расщепления предшествующей порции жиров, лецитина и желчных кислот жиры
образуют эмульсию с размером капелек около 5 нм. В тонком кишечнике жиры
стимулируют выделение клетками слизистой холецистокинина,
активирующего секрецию ферментов поджелудочной железы и сокращения желчного
пузыря.
Липаза, секретируемая
поджелудочной железой, состоит из двух компонентов –колипазы, образующейся в результате
активации проколипазы трипсином и локализованной на границе раздела водной и
липидной фаз, и липазы, образующей комплекс с колипазой. Липаза
катализирует отщепление от триглицеридов жирных кислот в положениях 1 и 3 с
образованием 2–моноглицеридов. Количество липазы, поступающей с
панкреатическим соком, так велико, что к тому моменту, когда жир достигает
середины двенадцатиперстной кишки, 80% его оказывается гидролизованным. В связи
с этим нарушение переваривания жиров, связанное с недостаточностью липазы, не
выявляется вплоть до полного прекращения деятельности поджелудочной железы или
сильного ее разрушения.
Помимо липазы поджелудочная
железа секретирует и другие ферменты липидного обмена, также активируемые
трипсином. К числу этих ферментов относится фосфолипаза А2, которая в
присутствии ионов Са2+ и желчных кислот отщепляет жирную кислоту от
фосфолипида лецитина с образованием лизолецитина. Холестерол
обычно присутствует в пище в виде эфиров и высвобождается под действием холестеролэстеразы.
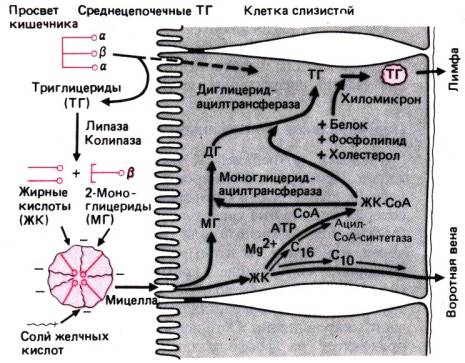
Рис. 29.38. Переваривание и всасывание липидов. В просвете
кишечника триглицериды расщепляются под действием колипазы и липазы до жирных
кислот и 2–моноглицеридов, которые содержатся в растворе в виде мицелл и
поступают из них в энтероциты. В клетках из длинноцепочечных жирных кислот и
2–моноглицеридов ресинтезируются триглицериды, которые в виде заключенных в
белковую оболочку хиломикронов выходят в лимфу. Жирные кислоты с короткими или
средними цепями поглощаются и переносятся в кровь непосредственно в этой форме.
МГ–моноглицериды, ДГ–диглицериды, ТГ–триглицериды, ЖК –жирные кислоты (по [27]
с изменениями)
Продукты гидролиза липидов плохо растворимы в воде и
могут находиться в кишечнике в растворенном виде лишь в составе мицелл.
Простые мицеллы, состоящие только из желчных кислот (чистые мицеллы), после
внедрения в их гидрофобную сердцевину жирных кислот, моноглицеридов,
фосфолипидов и холестерола превращаются в смешанные мицеллы. Благодаря
растворимости этих мицелл в воде концентрация конечных продуктов
гидролитического расщепления липидов в просвете кишечника повышается в тысячи
раз. Жирные кислоты с короткими и средними цепями и содержащие их липиды
довольно хорошо растворимы в воде и могут диффундировать к поверхности
энтероцитов, не встраиваясь в мицеллы.
Всасывание продуктов гидролитического расщепления
жиров. Жиры всасываются так
эффективно, что 95% триглицеридов (но лишь 20–50% холестерола) поглощается уже
из просвета двенадцатиперстной кишки и верхнего отдела тощей кишки. У человека
при обычной диете в сутки выделяется с калом до 5–7 г жира. При безжировой
диете эта величина уменьшается до 3 г/сут, а источником жира служат
слущивающиеся эпителиальные клетки и бактерии.
Прежде чем попасть внутрь энтероцита, компоненты
смешанных мицелл должны преодолеть три барьера:
1) неперемешивающийся
водный слой, прилежащий к поверхности клетки,–основное препятствие для
жирных кислот с длинными цепями и моноглицеридов и для выполнения мицеллами их
функций;
2) слой слизи, покрывающий
щеточную каемку; при толщине 2–4 мкм этот слой также препятствует переносу
компонентов мицелл;
3) липидную мембрану
энтероцита. Мицеллы в клетку не проникают, но их липидные компоненты
растворяются в плазматической мембране и быстро диффундируют в клетку по
концентрационному градиенту. Остаточное вещество мицелл может затем
возвратиться в просвет и включить новые липидные компоненты.
Внутриклеточный синтез липидов. Пройдя через клеточную поверхность, продукты
расщепления жиров – моноглицериды и жирные кислоты– транспортируются
небольшим белком с мол. массой 12000 в микросомы эндоплазматического
ретикулума, где из них вновь синтезируются триглицериды и другие липиды.
Вначале при участии моноглицеридтрансферазы образуются эфиры активированных
жирных кислот с моноглицеридами, а затем при участии диглицеридтрансферазы к
синтезированному диглицериду присоединяется еще одна жирная кислота и
образуется триглицерид. Другой, менее важный путь ресинтеза триглицеридов
связан с обменом глюкозы.
Фосфолипиды, как и триглицериды, могут образовываться в
энтероцитах путем эстерификации (примером служит образование лецитина из
лизолецитина). Реэтерификация холестерола катализируется холестеролэстеразой.
Кроме того, в подвздошной кишке происходит синтез холестерола de novo, поэтому тонкому кишечнику принадлежит особая роль
в обмене этого липида.
Образование хиломикронов. Новообразованные триглицериды и другие липиды,
прежде чем покинуть клетку, покрываются специальной оболочкой, содержащей холестерол
и фосфолипиды в сочетании со специфическими гликопротеинами, синтезированными в
аппарате Гольджи. Состав этих сложных образований, называемых хиломикронами,
приблизительно следующий: 90%–триглицериды, 7%–фосфолипиды, 2%–холестерол и
1%–белок. Диаметр хиломикронов составляет 60–75 нм в зависимости от скорости
всасывания и ресинтеза жиров.
Хиломикроны накапливаются в секреторных
везикулах; последние сливаются с боковой мембраной клетки, и через
образующееся при этом отверстие хиломикроны выходят в межклеточное
пространство, откуда по центральному лимфатическому и грудному
протокам поступают в кровь. После приема жирной пищи содержание
хиломикронов в плазме повышается так сильно, что она становится по цвету
молочно–белой (алиментарная гиперлипемия). Известно врожденное
заболевание–А–b–липопротеинемия,–связанное
с нарушением высвобождения хиломикронов из клеток. При этом заболевании
поглощенный жир не выводится из клетки и остается в ней. Помимо хиломикронов в
энтероцитах образуются липопротеины очень низкой плотности, которые
также высвобождаются в лимфатические сосуды.
Триглицериды, содержащие
короткоцепочечные и среднецепочечные жирные кислоты. В отличие –от триглицеридов,
содержащих жирные кислоты с длинной цепью, до 30% триглицеридов, содержащих
жирные кислоты с короткими и средними цепями, захватываются клетками в интактном
виде. Внутри клетки жирные кислоты отщепляются под действием эстераз и
вместе с жирными кислотами, поступившими в энтероциты в свободном виде,
диффундируют из клеток и поступают через капилляры в воротную вену.
Существование этого механизма, значительно более простого, чем механизм
транспорта триглицеридов, содержащих жирные кислоты с длинными цепями,
используют при лечении больных с нарушениями переваривания или всасывания
липидов – путем замены в диете триглицеридов с длинноцепочечными жирными
кислотами на триглицериды со среднецепочечными жирными кислотами.
Патофизиологические аспекты. Всасывание жиров в тонком
кишечнике может быть нарушено в связи с дефектами механизмов расщепления жиров
(неперевариваемость) или поглощения клетками продуктов расщепления (мальабсорбция).
Термин неусвояемость подразумевает оба вида расстройств. Часто причиной
неперевариваемости служит нарушение функции поджелудочной железы, а типичным
синдромом мальабсорбции–«брюшная болезнь», при которой вершины ворсинок
тонкого кишечника разрушаются вследствие гиперчувствительности к клейковине.
Симптоматика при обоих видах расстройств одинакова–потеря в весе, авитаминоз,
понос и жирный стул.
Дифференциальная диагностика проводится путем проб на
экзокринную функцию поджелудочной железы, различных видов обследования
(рентгенологического, эндоскопического) и биопсии.
29.7. Толстый кишечник
В толстом кишечнике химус перемешивается
под действием непропульсивной перистальтики, концентрируется в результате
обратного всасывания воды и подвергается дальнейшему расщеплению под действием
бактерий. Непереваренные остатки пищи под действием пропульсивной перистальтики
продвигаются в виде каловых масс по направлению к прямой кишке.
Функциональная
анатомия
Толстый кишечник человека
имеет в длину 120–150 см; его диаметр составляет 6–9 см в области слепой кишки
и уменьшается в дистальных отделах. Разные отделы толстого кишечника выполняют
специальные функции. В слепой кишке, где химус еще имеет жидкую консистенцию,
преобладает бактериальное расщепление и всасывание воды. Эти процессы
продолжаются в восходящей, поперечной и нисходящей кишках.
Продвигаясь по ним, содержимое кишечника приобретает все более плотную
консистенцию. Сигмовидная и прямая кишки служат главным образом
резервуарами. Толстый кишечник ограничен двумя сфинктерами – илеоцекалъной
(баугиниевой) заслонкой на проксимальном конце и анальным сфинктером
на дистальном. Последний состоит из внутреннего кольца гладких мышц и наружного
кольца поперечнополосатых мышц.
В проксимальном отделе
толстого кишечника продольный слой мышц состоит из трех полос (тений) по
0,8 см шириной. Благодаря тонусу этих мышц и локальным сокращениям циркулярных
мышц в толстом кишечнике образуются серии вздутий (гаустр) и перетяжек.
Начиная с сигмовидной кишки, продольные мышцы опять покрывают поверхность
кишечника сплошным слоем и, соединяясь в дистальном отделе, переходят в
мышечную оболочку прямой кишки. В слизистой оболочке толстого кишечника
отсутствуют ворсинки, подобные ворсинкам тонкого кишечника, но имеются микроворсинки.
Слизистая оболочка образует крипты глубиной 0,7 мм, выстланные эпителиальными
клетками, многочисленными слизистыми клетками и небольшим числом эндокринных
клеток. Клетки мигрируют со дна крипты к ее вершине и здесь слущиваются. Весь
процесс занимает 5–7 дней, т. е. несколько больше, чем в тонком кишечнике.
Собственная пластинка обильно снабжена лимфоидной тканью и плазматическими
клетками, особенно в области аппендикса, где она получила название
«кишечных миндалин». Обилие лимфоидной ткани в кишечнике имеет большое значение
в связи с присутствием здесь большой популяции бактерий. Иммунокомпетентные
клетки секретируют преимущественно иммуноглобулин IgA.
Моторика
толстого кишечника
Регуляция. Характер
и частота сокращений толстого кишечника определяются 1) изменениями
потенциала гладко мышечных клеток, т. е. медленными волнами, связанными с
их потенциалами действия, и 2) модулирующим воздействием вегетативной
нервной системы и желудочно–кишечных полипептидов.
Парасимпатические импульсы активируют сокращения толстого кишечника. Это
связано с тем, что ацетилхолин вызывает деполяризацию мембран мышечных клеток и
при достижении пороговой величины потенциала действия возникают медленные
волны. Симпатическая стимуляция и норадреналин, напротив, вызывают
гиперполяризацию, приводящую к расслаблению мышц.
Посредством этих эффектов вегетативной нервной
системы на моторику толстого кишечника может влиять центральная нервная
система. Это влияние можно продемонстрировать, подвергая субъекта «стрессовому
интервью» и одновременно измеряя с помощью чувствительного баллона давление
в толстой кишке. Когда разговор вызывает агрессивную реакцию, враждебность и
напряжение, давление в кишке повышается. Депрессия, страх, печаль, напротив,
вызывают снижение давления.
На моторику толстого кишечника могут также влиять
желудочно–кишечные полипептиды, которые либо усиливают (гастрин и холецистокинин),
либо угнетают ее (секретин, глюкагон). Частота медленных волн варьирует
в толстом кишечнике сильнее., чем в тонком, но при этом не существует градиента
в проксимально–дистальном направлении. На протяжении восходящего и нисходящего
отделов толстой кишки средняя частота волн составляет 6/мин; в слепой и
сигмовидной кишке она меньше, а в прямой кишке больше (17/мин). Нейроны
межмышечного нервного сплетения оказывают угнетающее действие на пейсмекеры
гладкомышечных клеток, и его отсутствие может приводить к сильным нарушениям,
как в случае болезни Гиршпрунга. При этом врожденном заболевании в
определенном участке толстого кишечника отсутствует нервное сплетение и участок
находится в состоянии постоянного тонического сокращения. Эвакуация содержимого
кишки задерживается, поэтому выше этого участка скапливаются каловые массы, в
результате чего толстый кишечник сильно растягивается. Это состояние называется
мегаколоном.
Наряду с
ингибирующими нейронами межмышечное нервное сплетение содержит стимулирующее
холинергические и пептидергические нервные волокна, медиаторами в которых
служат вещество Р, энкефалин и соматостатин.
Виды сократительной активности. При обычной для жителей развитых стран диете с
низким содержанием в пище грубоволокнистых веществ время продвижения химуса
от илеоцекальной заслонки до прямой кишки составляет 2–3 сут. Частички пищи,
находящиеся в центре химуса, могут проходить через толстый кишечник за более
короткое время. Время прохождения, равное 2–3 сут, было установлено
экспериментальным путем. Обследуемому давали вместе с пищей мелкие частицы
контрольного вещества (маркера) и фиксировали время, необходимое для того,
чтобы 80% маркера вышло с калом. При увеличении содержания в пище
грубоволокнистых компонентов время эвакуации может сокращаться при одновременном
увеличении массы кала. У сельских жителей Африки, потребляющих много
волокнистых веществ, среднее время эвакуации из толстого кишечника составляет
36 ч, а масса кала–480 г, тогда как у жителей европейских городов
соответствующие величины составляют 72 ч и 110 г. Большая продолжительность
эвакуации из толстого кишечника свидетельствует о том, что его моторика
является в основном непропульсивной. Сокращения циркулярных мышц не
имеют упорядоченного поступательного характера; они могут наблюдаться одновременно
в нескольких местах и служат скорее для перемешивания содержимого кишечника,
чем для его продвижения. При последовательном сокращении циркулярных мышц двух
соседних гаустр содержимое кишечника продвигается приблизительно на 10 см, но
при этом движение может происходить как в проксимальном, так и в дистальном
направлениях. В таком сокращении могут иногда участвовать более двух сегментов.
На простые гаустральные сокращения приходится более 90% всей моторики толстого
кишечника.
Изредка возникает истинная перистальтическая
волна, при которой сокращению предшествует расслабление; тогда содержимое
кишечника продвигается приблизительно на 20 см, причем продвижение может
происходить как в дистальном, так и в проксимальном направлении. Несколько раз
в день наблюдаются сильные сокращения–«перистальтические броски»,
получившие название феномена Холцкнехта по имени впервые описавшего их
рентгенолога. При этих сокращениях содержимое кишечника продвигается сразу на большое
расстояние, перемещаясь из поперечной кишки в сигмовидную.
Все упомянутые выше виды моторики могут усиливаться
после приема пищи. На моторную функцию кишечника влияют энергетический
потенциал и состав пищи, тогда как ее объем и рН не имеют значения. Калорийная
пища с высоким содержанием жиров усиливает моторику, а углеводы и белки на нее
не влияют. Усиление моторики начинается уже через 10 мин после приема пищи,
поэтому эффект называют желудочно–кишечным рефлексом. Он
предположительно включает холинергическую стимуляцию, поскольку
подавляется антагонистами ацетилхолина. По–видимому, в данном случае
ацетилхолин действует, вызывая высвобождение гастрина и холецистокинина.
Патофизиологические аспекты. Нарушения моторики толстого
кишечника приводят к запорам или поносам. Как следует из предшествующего
обсуждения, широко распространенное мнение о том, что усиление моторики
толстого кишечника вызывает только поносы, ошибочно. Может наблюдаться
противоположная картина, а именно что интенсификация непропульсивных
сокращений приводит к запорам. Примером могут служить спастические запоры,
которые часто сопровождают общее функциональное расстройство, называемое
«синдромом раздраженной толстой кишки». И наоборот, ослабление
сократительной активности толстого кишечника может вызывать понос в связи с
отсутствием резистентности, обусловленной в норме локальными сегментарными
сокращениями. С другой стороны, известны заболевания толстого кишечника
(например, язвенный колит, хроническое воспаление толстого кишечника), при
которых поносы связаны с более частыми массированными пропульсивными
сокращениями.
Переваривание и всасывание
Во время продвижения по
толстому кишечнику жидкий химус превращается в плотные каловые массы в
результате деятельности бактерий и всасывания воды.
Всасывание электролитов и воды. За день в слепую кишку вместе с химусом поступает
1–1,5 л жидкости. Около 90% этого объема всасывается в толстом кишечнике и
только около 100 мл выделяется с калом (рис. 29.33). При постепенном
поступлении жидкости может всосаться значительно больше воды (до 5 л), и только
после превышения этого объема (например, в результате секреторных процессов в
тонком кишечнике) наблюдается «диарея переполнения».
Электролиты и вода всасываются в толстой кишке
чрезвычайно эффективно даже против очень высокого осмотического градиента. Это
объясняется относительной плотностью эпителия толстого кишечника. Размер
пор плотных контактов в эпителии толстого кишечника составляет 0,23 нм. Из–за
высокого сопротивления мембраны вода поступает в межклеточное пространство
довольно медленно. Плотные контакты препятствуют обратному движению воды в
просвет кишечника, поэтому в толстом кишечнике поглощение ионов Na+
может продолжаться даже тогда, когда содержимое кишечника становится
гипотоническим, до концентрации Na+ 30 мМ. Плотность эпителия сочетается в толстом кишечнике с
высокой разностью потенциалов, создаваемой электрогенным транспортом Na+ у человека разность
потенциалов составляет 30–40 мВ в толстой кишке и только 2–4 мВ в тощей кишке.
Вдоль самого толстого
кишечника существует градиент «плотности» эпителия. В слепой кишке
эпителий наименее плотный и всасывает воду из жидкого химуса быстрее и в
большем объеме, а в прямой кишке эпителий имеет наибольшую плотность и
может всасывать ионы Na+ даже из относительно
плотного по консистенции содержимого кишечника, вследствие чего каловые массы
гипотоничны по отношению к плазме.
Ионы Na+ поступают в
клетки путем простой диффузии за счет большого градиента концентраций (130
ммоль/л) и разности потенциалов (—30 мВ) (рис. 29.39). Выведение Na+ из клеток осуществляется в
базолатеральной области клеточной мембраны насосом, активируемым (Nа+К+–АТФазой.
Ионы К+ выходят по электрохимическому
градиенту из межклеточного пространства в просвет кишечника через относительно
проницаемые для них плотные контакты. Несмотря на высокую концентрацию ионов К+
в клетке (около 80 мМ), лишь небольшая их часть диффундирует в просвет
кишечника непосредственно через клеточную мембрану. Различие в механизмах транспорта
ионов Na+ и К+ отражается на их соотношении в
каловых массах. Ежедневно в толстый кишечник поступает 5–10 ммоль К+,
а выводится с каловыми массами 10–15 ммоль, при этом концентрация в последних
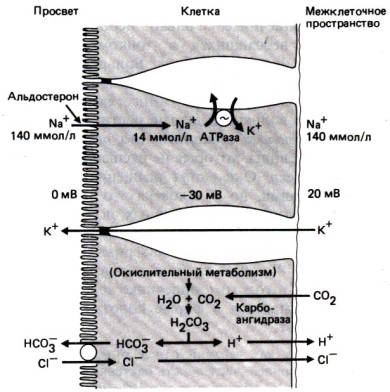
Рис. 29.39. Транспорт ионов в слизистой
толстого кишечника. Вверху: активное поглощение Na+ (стимулируется альдостероном) и пассивная диффузия К+
в противоположном направлении. Внизу: обмен ионов НСО3–
и Сl–
(по [1])
К+ составляет примерно 90 ммоль/л. С другой
стороны, в слепую кишку поступает 150 ммоль Na+, и его концентрация здесь
составляет 130 ммоль/л, а экскретируется только 2–4 ммоль ионов Na+, и концентрация их в
каловых массах составляет лишь 40 ммоль/л.
Поглощение Na+ и выделение К+
возрастают под действием минералокортикоидов, когда концентрация
последних в плазме повышается в физиологических пределах (как это имеет место
при потере натрия), и глюкокортикоидов, вводимых в фармакологических
количествах при лечении хронических воспалительных процессов. Эти гормоны
стимулируют (Na+–K+)–ATФaзy, а альдостерон, кроме того,
повышает проницаемость апикальной области клеточной мембраны для Na+.
Хлорид и бикарбонат также активно всасываются в
толстом кишечнике. Ежедневно в толстый кишечник поступает около 60 ммоль ионов
С1–, и его концентрация здесь составляет около 60 мМ, но с калом
выводится только около 2 ммоль ионов С1–, а концентрация их в кале
не превышает 15 мМ. Низкая конечная концентрация ионов С1– позволяет
предположить, что существует их активный транспорт против высокого
химического градиента. Поглощение С1– сопряжено с выделением НСO3– в обменной системе (рис.
29.39). Благодаря накоплению ионов НСО3– в просвете
кишечника каловые массы имеют слабощелочную реакцию.
Помимо воды и ионов в
толстый кишечник поступают органические вещества, не всосавшиеся в тонком
кишечнике. Только небольшая часть веществ, не всосавшихся в тонком кишечнике,
вообще способна всасываться. Большая же часть этих веществ не всасывается,
поскольку они не перевариваются. К таким веществам относятся растительные
волокна и другие растительные компоненты – целлюлоза, гемицеллюлоза, пектины
и лигнины, т. е. углеводы, которые не расщепляются амилазами человека.
Они могут расщепляться кишечными бактериями, но у человека это
расщепление идет не так активно, как у жвачных, получающих за счет
бактериального расщепления углеводов основную массу энергетического материала.
И тем не менее около 40–50% грубоволокнистых веществ расщепляется в толстом
кишечнике человека бактериями и всасывается в форме короткоцепочечных жирных
кислот.
В то время как верхний отдел
тонкого кишечника практически стерилен, численность бактерий вдоль кишечника
возрастает. Особенно резко (в 104 –105 раз) увеличивается
популяция бактерий за пределами илеоцекальной заслонки, где она составляет 1011
–1012 против 106 мл химуса в подвздошной кишке. Большая
часть бактерий толстого кишечника–строгие анаэробы Bifidus и Bacteroides (грамположительные и грамотрицательные
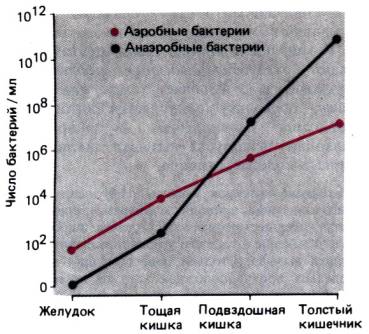
Рис. 29.40. Популяция бактерий желудочно–кишечного тракта. В
желудке и тощей кишке преобладает аэробная флора ротовой полости. При переходе
из подвздошной кишки в слепую происходит заметное увеличение популяции,
особенно численности анаэробных бактерий (обратите внимание на логарифмический
характер оси ординат) (по [2])
бактерии, не образующие
спор). Аэробные бактерии –Е. coli, энтерококки и лактобактерии
составляют менее 1% от общего числа бактерий толстого кишечника. В толстом
кишечнике присутствует более 400 видов бактерий, составляющих 30– 50% (по
другим расчетам 75%) сухого вещества каловых масс [42].
Анаэробные бактерии
расщепляют растительные волокна до жирных кислот, состоящих из 2–4
углеродных атомов (уксусной, масляной и пропионовой). Эти кислоты дают 50%
анионов, присутствующих в толстом кишечнике. Большая часть анионов
нейтрализуется бикарбонатом.
Хотя белки, углеводы, жирные
кислоты с длинной цепью и глицериды в толстом кишечнике не всасываются, жирные
кислоты с короткой цепью всасываются путем пассивной неионной диффузии.
Источником жирных кислот с длинной цепью служат непереваренные или
невсосавшиеся жиры пищи, и их количество увеличивается при нарушении
переваривания или всасывания жиров в тонком кишечнике. Эти жиры могут также
расщепляться бактериальными липазами, и около 25% образующихся при этом жирных
кислот с длинной цепью подвергается гидроксилированию под действием бактерий.
Гидроксилированные жирные кислоты вызывают понос подобно рициновой кислоте –
активному компоненту касторового масла.
Дефекация
и диарея
Поступление каловых масс в
прямую кишку вызывает рефлекторный акт дефекации. У жителей западных стран
нормальная частота актов дефекации колеблется от 3 раз в сутки до 3 раз в
неделю. Частота актов дефекации сильно зависит от состояния толстого кишечника,
и в первую очередь от его моторики и содержания воды в каловых массах.
Поскольку диарея наблюдается при увеличении содержания воды в кишечнике
до 200–300 мл, ее может вызвать уже небольшое нарушение всасывания воды в
толстом кишечнике.
Патофизиологические
аспекты. При дефиците лактазы в
тонком кишечнике (интолерантности к молоку) лактоза не расщепляется и в большом
количестве поступает в толстый кишечник. Некоторая часть лактозы расщепляется в
толстом кишечнике бактериями, но оставшегося количества достаточно для того,
чтобы вызвать приток воды и понос, поскольку лактоза обладает большой
осмотической активностью. Тот же эффект лежит в основе послабляющего действия
таких плохо поглощающихся ионов, как ионы магния или сульфата. Все это примеры осмотической
диареи.
Другие группы слабительных действуют за счет
превращения плотных контактов в «протекающие» контакты и изменения таким
образом всасывающих свойств эпителия толстого кишечника. Слизистая толстого
кишечника напоминает в этих случаях слизистую тонкого кишечника, так как
неспособна поддерживать высокие градиенты, необходимые для задержки ионов Na+
и воды. К таким веществам относятся гидроксилированные жирные кислоты, в
том числе рициновая кислота, желчные кислоты, не подвергшиеся всасыванию
в тонком кишечнике и достигшие толстого кишечника в достаточно высокой
концентрации (хологенная диарея), а также дифенольные слабительные типа
бисакодила. Некоторые из этих слабительных подавляют также транспорт ионов Na+ за счет угнетения (Na+–K+)–АТФазы.
29.8. Литература
Учебники и руководства
1.
Clinical gastrointestinal physiology. Granger D. N.. Barrowman J. A.,
Kvietys P. R., eds. W. B. Saunders Philadelphia, 1985.
2. Colon, structure and function. L. Bustos–Fernandes ed. Plenum
Medical Book Сотр. New York, 1983.
3. Digestion and the structure and function of the gut. Magee D.F..
Dalley A.F.. eds. Karger Basel, 1986.
4. Dumdarm A. Handbuch Innere Medizin, 5. Aufl. Bd. 3,
Verdauungsorgane Teil 3, A. Caspary W.F., ed. Springer Berlin, 1983.
5. Gastrointestinal disease: pathophysiology, diagnosis, management. Sleisenger
M. H., Fordtran J. S., eds. W. B. Saunders Co., Philadelphia, 1983.
6. Gastrointestinal pathophysiology. Brooks F. P., ed. Oxford
Univ. Press, New York, 1974.
7. Gastrointestinale Physiologic. Konturek S. J., Ciassen M., eds.
G. Witzstock Baden–Baden, 1976.
8. Handbook of Physiology. Section 6: Alimentary Canal. Vol. 1–5. Code
C. ed. Am Physiol. Soc. Washington, 1968.
9. Intestinal absorption and
secretion. Scadhauge E.,
Heintze K., eds. MTP Press Lim. Lancester, 1984.
10. Medical physiology. Mountcastle V. В., ed. C. V. Mosby Co. St. Louis, 1980.
11. Physiology of the digestive tract. Davenport H. W., 5th
edition. Year Book Med. Publ. Chicago, 1982.
12. Physiology of the gastrointestinal tract. Johnson L. R., ed.
Raven Press, New York, 1981.
13. Undergraduate teaching project in gastroenterology and liver
disease. American Gastroenterological Association Timonium. Milner–Fernwick,
I.N.C., 1976.
14. Taschenatlas der Physiologic. Silbernagi S., Despopulos A. Thieme
Stuttgart, 1979.
15. The large intestine. Wrong O.M., Edmonds C.J., Chadwick V.S.,
eds. MTP Press Lim. Lancester, 1981.
Оригинальные статьи и обзоры
16. Anderson
D.J„ Mastication. In: Handbook of Physiology, Section 6. Alimentary canal.
Vol. 4, Motility. Code C ed. Washington. Am. Physiol. Soc., 1968, pp.
1811–1820.
17. Claude P., Goodenough D. A. Fracture faces of zonula
occludentes from "tight" and "leaky" epithelia. J. Cell.
Biol., 58, 390–400 (1973).
18. Eckhardt R., Meyer zum Buschenfelde K.H. Immunologie des
Diinndanns. In: Handbuch Innere Medizin 5. Aufl. Bd. 3; Verdauungsorgane. Teil
3 A. Caspary W. F., ed. Springer Verlag, 1983, S. 73–104.
19. Elsenhans В., Caspary W.F.
Resorption von Kohlenhydraten. In: Handbuch Innere Medizin 5. Aufl., Bd. 3;
Verdauungsorgane, Teil 3 A. Caspary W.F., ed. Springer Berlin, 1983, S.
139–156.
20. Elsenhans В., Caspary W.F.
Resorption von Eiweiss. In:
Handbuch Innere Medizin 5. Aufl., Bd. 3; Verdauungsorgane, Teil 3 A. Caspary
W.F., ed. Springer Berlin, 1983, S.157–178.
21. Fellenius E., Berghlind Т.,
Sachs G., Olbe L., Elander B. Sjostrand S. E., Wallmark B. Substituted benzimidazoles inhibit acid secretion by blocking (Na+K+–ATPase, Nature, 290, 159–161 (1981).
22. Fordtran J. S., Locklear T. W. Ionic constituents and
osmolality of gastric and small intestinal fluids after eating. Am. J. Dig.
Dis., 11, 503–521 (1966).
23. Forth W. Intestinale Resorbtion von Eisen und chemisch
verwandten Metallen. In: Handbuch Innere Medizin 5. Aufl. Bd. 3;
Verdauungsorgane, Teil 3 A. Caspary W.F., ed. Springer Berlin, 1983, S.
267–297.
24. Fromter E., Diamond J. Route of passive ion–permeation in epithelia.
Nature, 235, 9–13 (1972).
25. Fromm H., Bazzoli F. Enterohepatischer Kreislauf der
Gallensaure. In: Handbuch Innere Medizin 5. Aufl., Bd. 3;
Verdauungsorgane Teil 3 A. Caspary W.F., ed. Springer Berlin,
1983, S. 352–372.
26. Gang! A. Resorption von Triglyceriden und fettloslichen
Vitaminen (ausser Vitamin D). In: Handbuch 1'nnere Medizin 5. Aufl., Bd. 3;
Verdauungsorgane Teil 3 A. Caspary W.F., ed. Springer Berlin 1983, S.
179–215.
27. Gray G. M. Mechanisms of digestion and absorption of food.
In: Gastrointestinal disease. 2nd ed. Sleisenger M. H., Fortran S.J.,
eds. Philadelphia, W.B. Saunders Co., 1983.
28. Hentschel E. Cimetidin. In: Ulcustherapie. Blum A. L.,
Sievert J.R.. eds. Springer Berlin, 1982, S. 225–253.
29. Hofmann A. F. The enterohepatic circulation of bile acids in
health and disease. In: Gastrointestinal disease. Sleisenger M.H.. Fordtran
J.S., eds. W. B. Saunders, Philadelphia, 1983. p.115–132.
30. Ito S. Functional gastric morphology. In: Physiology of the
gastrointestinal tract. Johnson L. R., ed. Raven Press New York, 1981,
p. 517–550.
31. Kreijs G.J. Wasser– und Elektrolyttransport des Diinndarms.
In: Handbuch Innere Medizin 5. Aufl., Bd. 3;Verdauungsorgane, Teil 3 A. Caspary
W.F., ed. Springer Berlin, 1983, S. 434–463,
32. Lembcke В., Caspary W. F.
Intestinale Gasproduction. In: Handbuch Innere Medizin 5. Aufl., Bd. 3;
_Verdauungsorgane Teil 3 A. Caspary W.F., ed. Springer Berlin, 1983, S.
521–541.
33. Levitt M. D., Bond J. H., Levin D. G. Gastrointestinal gas.
In: Physiology of the gastrointestinal tract. Johnson L. R., ed. Raven
Press, New York, 1981, p. 1301–1316.
34. Nell G. Resorption von Gallensauren. In: Handbuch Innere
Medizin 5. Aufl., Bd; Verdauungsorgane Teil 3 A. Caspary W.F., ed.
Springer Berlin, 1983, S. 337–349.
35. Peerenboom H. Resorption von Kalzium, Magnesium und Phosphat
und ihre Regulation. In: Handbuch Innere Medizin 5. Aufl., Bd. 3;
Verdauungsorgane Teil 3 A. Caspary W.F., ed. Springer Berlin, 1983, S.
233–266.
36. Polak J. M., Bloom S. R., Sullivan S. N.. Frazer P., Pearse
A.G.E. Enkephaline–like immuno reactivity in the human gastrointestinal
tract. Lancet 1, 972–974, 1977.
37. Rumberger E. Physiologie des Colons. In: Handbuch Innere
Medizin 5. Aufl., Bd. 3; Verdauungsorgane Teil 4, Mulier–Weland K., ed.
Springer Berlin, 1982.
38. Rummel W. Biologische Mcmbranfunktion. Wirkungen von
Gallensauren und Laxantien auf den mukosalen Transfer. Cat. Med. Wiss, 32,
233–250 (1976).
39. Ruppin H. Motilitat des Dunndarms. In: Handbuch Innere
Medizin 5. Aufl., Bd. 3; Verdauungsorgane Teil 3 A.Caspary W.F., ed.
Springer Berlin, 1983, S. 464–487.
40. Schiller L. R. Motor function of the stomach. In:
Gastrointestinal disease. Sleisenger M. H., Fordtran J. S., eds. W.B.
Saunders Philadelphia, 1983, p. 521–541.
41. Schullz S.G. Some properties and consequencies of
low–resistance paracellular pathway across the small intestine:
the advantage of being "leaky". In; Intestinal permeation. Kramer
M., Lauterbach F„ eds. Amsterdam. Excerpta medica, 1977, pp. 321–392.
42. Stephen A. M., Cummings J. H. The microbial contribution to
human faecal mass. J. Med. Microbiol., 13, 45–66 (1980).
43. Thaysen J.H., Thorn N. A., Schwartz I.L. Excretion of sodium,
potassium, chloride and carbon dioxide in human parotid saliva. Am. J. Physiol,
178, 155–159 (1954).
44. Trier J. S., Madara J. L. Functional morphology of the mucosa
of the small intestine. In: Physiology of the gastrointestinal tract. Johnson
L. R., ed. Raven Press New York, 1981, p. 925–962.
45. Weishrod N. W. Pattern of intestinal motility. Annual Rev.
Physiol, 43, 33–51 (1981).