Глава 24
ЭНЕРГЕТИЧЕСКИЙ БАЛАНС
Х.–Ф. Ульмер
24.1. Энергетический обмен
Энергетический обмен присущ каждой живой клетке;
богатые энергией питательные вещества усваиваются и химически преобразуются, а
конечные продукты обмена веществ с более низким содержанием энергии удаляются
из клетки. Освобождающаяся при этом энергия используется для различных целей,
например для поддержания клеточной структуры (и, следовательно,
сохранения ее функций}, а также для обеспечения специфических
клеточных активностей (таких, например, как сокращение мышечных клеток).
Обменные, или метаболические, процессы, в ходе
которых специфические элементы организма синтезируются из поглощенных пищевых
продуктов, называют анаболизмом; соответственно те метаболические
процессы, в ходе которых структурные элементы организма или поглощенные пищевые
продукты подвергаются распаду, называют катаболизмом. Метаболизм жиров и
углеводов служит главным образом для обеспечения физиологических функций
(функциональный метаболизм), тогда как белковый обмен нужен в первую
очередь для поддержания и изменения структуры организма (структурный
метаболизм).
Валовая продукция энергии. Интенсивность
энергопродукции организма в целом зависит от количества выделенной энергии
(внешняя работа, тепло) и от количества запасенной энергии (депонирование
питательных веществ, структурные преобразования) в единицу времени: общее
количество выработанной энергии–это сумма внешней работы, тепловых потерь и
запасенной энергии.
Единицы измерения
энергетического обмена. Традиционно энергетический обмен выражают в
килокалориях (ккал) на единицу времени. Однако в Международной системе единиц в
качестве основной единицы энергии принят джоуль (Дж): 1 джоуль = 1 ватт·1
секунда = 2,39–10–4 ккал; 1 ккал =4187 Дж = 4,187 кДж ≈ 0,0042
МДж. Отсюда следует, что 1 кДж/ч ≈ 0,28 Вт (≈ 0,239 ккал/ч) и 1
кДж/сут ≈ 0,012 Вт (≈0,239 ккал/сут).

Рис. 24.1. Практический коэффициент
полезного действия при различных видах физической работы (по Мюллеру, см. [7])
Коэффициент полезного действия. Если клетка совершает внешнюю работу, то часть
вырабатываемой при этом энергии обязательно выделяется в виде тепла (второй
закон термодинамики). Коэффициент полезного действия (η) активно
функционирующей клетки, как и КПД машины, представляет собой ту часть
вырабатываемой энергии, которая затрачивается на внешнюю работу; его величина
всегда меньше 100%:
η (%) =
Внешняя работа/ Вырабатываемая энергия •100 (1)
Следует различать суммарный коэффициент полезного
действия, рассчитываемый по общей знергопродукции, и практический
коэффициент полезного действия, определяемый по количеству выработанной
энергии за вычетом энергии основного метаболизма. Коэффициент полезного
действия изолированной мышцы в лучшем случае достигает 35%; при мышечной работе
целого организма его величина редко превышает 25% (см. примеры на рис. 24.1).
24.2. Параметры обмена
веществ
Параметры
обмена веществ в клетках
В связи с разнообразием метаболических функций в
живой клетке полезно выделить три основных уровня метаболической активности:
уровень активного обмена–интенсивность обменных процессов в активно
функционирующей клетке (изменяется в соответствии со степенью активности в
данный момент времени);
– уровень готовности – интенсивность
метаболизма, которую неактивная в данный момент клетка должна поддерживать для
того, чтобы сохранять способность к немедленному и неограниченному
функционированию–, этот уровень характерен, например, для процессов
поддержания определенной разности концентраций ионов Na+ и К+
– уровень поддержания – минимальная интенсивность метаболизма,
достаточная для сохранения клеточной структуры; если эта необходимая
потребность не удовлетворяется, в клетке происходят необратимые нарушения и она
погибает.
Данную классификацию уровней метаболизма необходимо
учитывать при оценке влияния нарушений энергетического обмена на
отдельную клетку или орган. Нарушения метаболизма могут быть вызваны различными
причинами, например уменьшением доставки кислорода или скорости кровотока либо
отравлением.
Уровень обмена веществ для организма в целом имеет
иное значение, нежели для отдельного органа. Так, например, если в дыхательной
мускулатуре или сердечной мышце метаболизм снизится до уровня готовности, эти
органы окажутся неактивными. В результате погибнут все клетки, поскольку
организм как целое не может выжить при бездействии дыхательных мышц или сердца.
Прекращение доставки энергии не вызывает немедленного
нарушения клеточной активности, поскольку в клетках имеются некоторые
энергетические резервы. Однако продолжительность времени, в течение которого
клетки могут сохранять свою функциональную активность в полном объеме, в
значительной степени зависит от того, какому органу они принадлежат. Если
органом, лишенным притока энергии, оказался головной мозг (например, в
результате полной ишемии), то примерно через 10 с наступает потеря сознания,
а через 3–8 мин в клетках возникают необратимые нарушения; однако в
том случае, если ишемии подвергается скелетная мышца, пребывающая в
состоянии покоя, интенсивность протекающих в ней обменных процессов не
становится ниже уровня поддержания на протяжении 1–2 ч.
Параметры обмена веществ в целом организме
Интенсивность обмена веществ
в условиях покоя.
Интенсивность метаболизма в покое не выражается
суммой соответствующих уровней готовности, присущих всем его клеткам, поскольку
некоторые органы (например, мозг, сердце, дыхательная мускулатура, печень и
почки) постоянно находятся в активном состоянии.
Интенсивность обмена веществ в организме в условиях
умственного и физического покоя нельзя оценить как строго определенное
численное значение, поскольку она подвержена влиянию различных факторов. Для
того чтобы легче было сравнивать результаты измерений интенсивности основного
обмена, установлены специальные условия проведения таких измерений.
Интенсивность основного обмена (ИОО). Обычно предусматриваются
следующие четыре условия для измерения интенсивности основного обмена: 1)
утром, 2) в покое (в лежачем положении), 3) натощак и 4) в условиях
температурного комфорта.
Интенсивность обмена веществ, измеренная утром,
натощак, в условиях покоя и температурного комфорта, когда–то служила
важным диагностическим показателем, который использовали для выявления
нарушений в функциях щитовидной железы. Впоследствии функциональное состояние
щитовидной железы стали определять другими способами, например с помощью
радиоактивного иода или путем измерения концентрации тиреоидного гормона в
крови. В настоящее время такой критерий, как интенсивность обмена веществ,
редко используют в диагностических целях.
Четыре стандартных условия измерения основного
обмена приняты с учетом следующих факторов, способных влиять на
интенсивность процессов обмена веществ у человека.
1. Интенсивность процессов обмена подвергается
суточным колебаниям – возрастает утром и снижается в ночной период.
2. Интенсивность процессов обмена возрастает в
условиях физической и умственной нагрузки, что связано с
увеличением числа клеток, интенсивность метаболизма в которых превышает уровень
готовности. В обоих указанных случаях основным органом, определяющим
интенсивность обмена веществ, являются мышцы (см. раздел, посвященный
интенсивности обмена веществ в условиях нагрузки, и рис. 24.2).
3. Интенсивность процессов обмена повышается во
время приема пищи и ее последующего переваривания, особенно если пища
была белковой. Этот эффект называют специфическим динамическим действием
пищи. Возрастание интенсивности метаболизма после еды связано не только с
пищеварительной активностью, но и с последующими процессами обмена веществ; оно
может продолжаться в течение 12ч, а в случае потребления большого количества белка
этот период может достигать 18 ч.
4. Интенсивность обмена веществ возрастает, если
температура окружающей среды отклоняется от комфортной (выходит за пределы диапазона
нейтральной температуры, или термонейтральной зоны; см. далее);
сдвиги в сторону охлаждения приводят к большему
усилению обмена веществ, чем сдвиги в сторону повышения температуры. Процессы
изменений температуры тела подчиняются закону Вант–Гоффа.
Интенсивность основного обмена наполовину
обусловлена метаболизмом печени и покоящейся скелетной мускулатуры (табл. 24.1). В связи с тем
что во время сна мышечный тонус снижается, интенсивность обмена веществ у
спящего или находящегося в состоянии наркоза человека может оказаться ниже
основного уровня. В условиях голодания интенсивность процессов обмена также
может падать ниже стандартного значения вследствие ослабления работы печени.
Нормальные величины основного обмена. Значения интенсивности
основного обмена у здоровых людей, измеренные даже в строго стандартных
условиях, варьируют. Вариабельность связана с влиянием таких факторов, как возраст,
пол, рост и масса тела (рис. 24.3).
Эти переменные учитываются в таблицах показателей
основного обмена, составленных на основании данных, полученных на больших
группах людей такими авторами, как Бутби и др. [10], Бутби и Дюбуа (см. [9]),
Флейш (см. [9]), Харрис и Бенедикт [13], а также Кестлер и Книппинг [2].
Небольшие различия между нормальными показателями
основного обмена, приводимыми в разных таблицах, частично связаны с тем, что
данные были получены для разных регионов (страны Северной Америки, Швейцария,
Германия). Средние и «нормальные» показатели всегда зависят от того, в какой
группе испытуемых проводится исследование. Как правило, в качестве примерного
значения
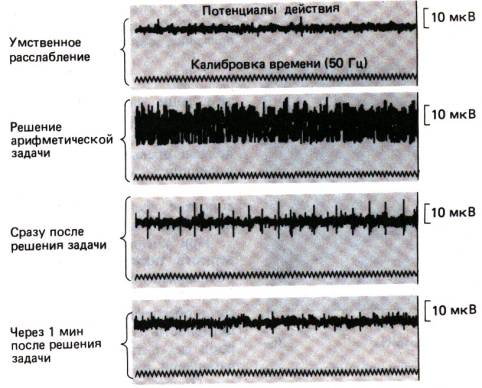
Рис. 24.2. Рефлекторное усиление
мышечного тонуса в период умственной работы, отраженное в записи потенциалов
действия (электромиограмма) мышцы предплечья (по [12] с изменениями)
Таблица 24.1. Относительный вклад
различных органов в обеспечение основного обмена у человека [4]
|
Орган:
|
Печень
|
Мышцы
|
Мозг
|
Сердце
|
Почки
|
Другие органы
|
|
Относительный вклад
|
26%
|
26%
|
18%
|
9%
|
7%
|
14%
|
стандартной интенсивности обмена веществ у взрослого
человека может быть принята величина, равная 4,2 кДж·кг–1 •ч–1
(1,2Вт); для человека, весящего 70 кг, соответствующий показатель основного
обмена составляет приблизительно 7100 кДж/сутки (84 Вт).
Интенсивность обмена веществ в условиях нагрузки. При физической
работе скорость метаболизма возрастает в зависимости от степени физического
напряжения. Интенсивность обмена веществ при «относительном покое», т.
е. у испытуемого в очень малоактивном состоянии, составляет приблизительно 8400
кДж/сут (97 Вт) для женщин и 9600 кДж/сут (110 Вт) для мужчин. Эта
величина соответствует суточному обмену веществ у значительной части
населения– людей, занимающихся «сидячей работой» и не затрачивающих
сколько–нибудь значительных физических усилий.
Применяемый в литературе термин «скорость
рабочего метаболизма» подразумевает интенсивность расхода энергии во
время физической работы (т.е. суммарные энергетические затраты для
поддержания основного обмена веществ и обеспечения физической деятельности).
Чем интенсивнее работа,
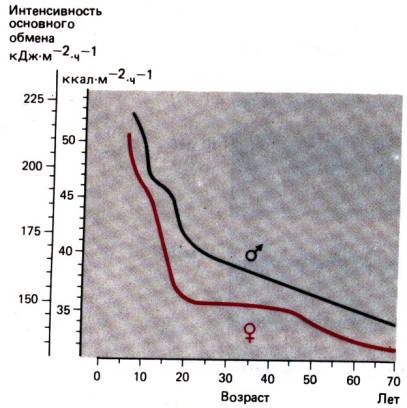
Рис. 24.3. Зависимость относительной
интенсивности основного обмена от возраста и пола. Площадь поверхности тела (м2)
является функцией роста и массы индивида [10]
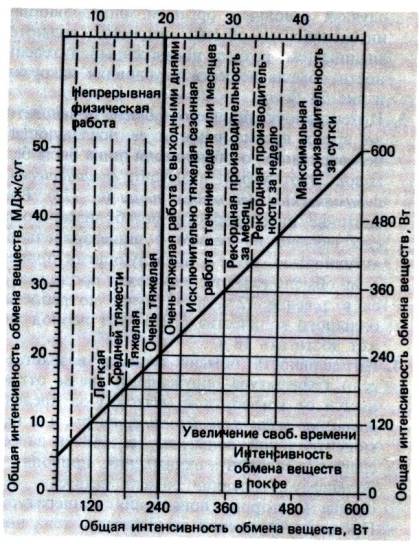
Рис. 24.4. Энергетические затраты (за сутки)
у мужчин при работах, требующих различного физического напряжения, и
классификация таких работ [4]
тем выше интенсивность рабочего метаболизма. У
людей, занятых на физической работе в течение многих лет, верхний предел
энергетических затрат не должен превышать 15 500 кДж/сут (около 186 Вт) у
женщин и 20 100 кДж/сут (около 240 Вт) у мужчин. Эти показатели могут быть
превышены лишь на короткое время или в том случае, если работа прерывается
несколькими днями отдыха (рис. 24.4).
При умственной работе интенсивность обмена
веществ также увеличивается, хотя вряд ли это обусловлено потребностями мозга.
Во время умственной работы сосредоточение на каком–либо предмете обеспечивается
просто за счет перемещения активности из одной области в другую; даже в период
сна в ткани мозга не происходит заметных изменений интенсивности метаболических
процессов. Причиной увеличения обмена веществ в период умственной работы служит
рефлекторное увеличение мышечного тонуса (рис. 24.2).

Рис. 24.5. Энергетические затраты (за
сутки) при работах разного рода и занятиях в свободное время (по [4] с
изменениями). Фактические значения могут различаться в зависимости от того, как
выполнялась работа
24.3. Интенсивность обмена веществ в особых условиях
На рис. 24.4 и в табл. 24.2 представлены значения
интенсивности обмена веществ при различных нагрузках. Интенсивность обмена при
работах разного рода и во время отдыха можно оценить по рис. 24.5 или пользуясь
подробными таблицами, опубликованными Спицером с сотр. [б]. Степень напряжения
при разных видах физической работы указана на рис. 24.4.
У спортсменов интенсивность обмена веществ может
возрастать в относительно более высокой степени, чем у работающих людей, но на
значительно более короткое время. Роль фактора времени проиллюстрирована на
рис. 24.4; особенно показательны величины интенсивности обмена при беге на
различные дистанции–от стометровки до марафона. Чем длиннее дистанция
(и, следовательно, больше затрачиваемое время), тем ниже уровень метаболизма
(рис. 24.6). Интенсивность обмена веществ при забеге на 100 или 200 м
составляет 22 кВт, что приблизительно в 13 раз больше, чем при беге на
марафонскую дистанцию. Работа, совершаемая за 10с бега с высокой
скоростью,
Таблица 24.2. Интенсивность обмена веществ
при определенных условиях (округленные значения) и соответствующие величины
поглощения кислорода (см. непрямое измерение интенсивности метаболизма)
|
Условия
|
Интенсивность метаболизма
|
Поглощение О2, мл/мин
|
|
МДж/сут
|
Вт
|
|
ИОО (интенсивность основного обмена.) при массе тела 70 кг
|
♀ 6,3
|
76
|
215
|
|
|
♂ 7,1
|
85
|
245
|
|
ИОО плюс интенсивность метаболизма
|
♀ 8,4
|
100
|
275
|
|
при относительном покое
|
♂ 9,6
|
115
|
330
|
|
«Рабочий» метаболизм (при очень тяжелой
|
♀ 15,5
|
186
|
535
|
|
работе на протяжении ряда лет)
|
♂ 20,1
|
240
|
690
|
|
Общая интенсивность метаболизма при занятиях
спортом (виды, требующие выносливости; результаты
выше средних)
|
кДж/ч
4300
|
Вт
1200
|
мл/мин
3600
|
1
.
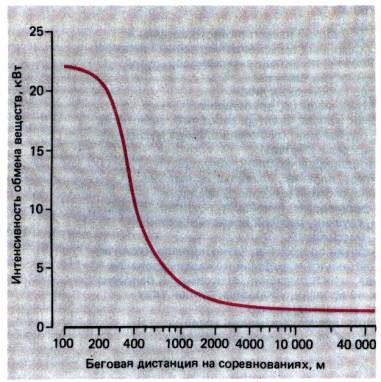
Рис. 24.6. Энергетические затраты при
соревнованиях по бегу на разные дистанции [6]
достигает 200 кДж, что соответствует величине
калорийности примерно 14 г глюкозы. За два с лишним часа, требующиеся для
преодоления марафонской дистанции, бегун затрачивает около 1,6 кВт, что
значительно больше приведенного на рис. 24.4 максимального дневного уровня
метаболизма при работе в течение нескольких дней. Интенсивность обмена веществ
у бегунов на марафонскую
Таблица 24.3. Интенсивность обмена веществ
при занятиях различными видами спорта (данные по бегу см. на рис. 24.6) [6]
|
Вид спортивных занятий
|
|
Вт
|
|
Езда на велосипеде по ровной местности
|
20 км/ч
|
545
|
|
Велосипедные гонки
|
40 км/ч
|
1735
|
|
Игра в футбол
|
|
790–1040
|
|
Игра в гандбол
|
|
885
|
|
Игра в волейбол
|
|
380–640
|
|
Плавание брассом
|
28 м/мин
|
460
|
|
Плавание брассом в одежде
|
28 м/мин
|
730
|
|
Соревнования по гребле
|
|
1715
|
|
Лыжный спорт:
|
|
биатлон
|
|
610
|
|
слалом
|
|
1605
|
|
скоростной спуск
|
|
|
|
женщины
|
|
2130
|
|
мужчины
|
|
3100
|
|
бег по пересеченной местности
|
|
женщины
|
|
1285
|
|
мужчины
|
|
1435
|
|
Теннис, одиночки
|
|
490– 1100
|
|
Танцы, венский вальс
|
|
335
|
дистанцию соответствует 2,1 «лошадиной силы» (750 Вт
= 1 л. с.). Если принять, что расщепляются примерно одинаковые количества жиров
и углеводов, то за 130–минутный марафонский пробег будет использовано 850 г
энергосодержащих питательных веществ. Величины интенсивности обмена веществ при
занятиях разными видами спорта приведены в табл. 24.3.
24.4. Методы измерения
Методы, с помощью которых измеряют интенсивность
метаболизма, можно классифицировать несколькими способами–по тому, заключаются
ли они в непосредственном измерении потерь тепла или в их измерении косвенным
путем, представляют ли собой используемые для измерения устройства открытые или
закрытые системы, переносные или стационарные, обеспечивают ли приборы
непрерывную или периодическую регистрацию интенсивности обмена
веществ.
Прямое измерение интенсивности обмена веществ
Данный метод основан на прямом определении тепловых
потерь организма («прямая калориметрия»). Еще в 1780 г. Лавуазье разработал
способ измерения тепла, выделяемого живыми организмами. Его «калориметр»
регистрировал количество выделяемого тепла прямым непрерывным способом, хотя и
не в стандартных условиях. Аппараты, необходимые для прямого измерения потерь
тепла у человека, массивны и сложны по устройству; в связи с этим их используют
только для специальных целей. Прямые измерения интенсивности обмена веществ
оказались особенно полезными в одном отношении: сравнение полученных этим
способом результатов с данными, характеризующими баланс пищевых веществ в
организме, подтвердило справедливость закона сохранения энергии для живых
организмов. Кроме того, результаты экспериментов по прямой калориметрии
могут быть полезны для контроля достоверности результатов, полученных
соответствующими непрямыми методами измерения.
Непрямое измерение
интенсивности обмена веществ
Непрямое измерение интенсивности обмена веществ
основано на измерении количества кислорода, поглощенного организмом. В
связи с тем что кислород используется в любой реакции биологического окисления,
а возможности хранения О2 в организме невелики, количество
кислорода, потребляемого тканями (показатель интенсивности метаболизма), можно
определить по количеству кислорода, посту пившего в организм через легкие.
Были предприняты также попытки использовать в
качестве меры количества вырабатываемого тепла количество выдыхаемого СО2.
Поскольку, однако, способности организма к накоплению углекислого газа весьма
велики, нет уверенности в том, что количество выдыхаемого СО2 достаточно
точно соответствует тому количеству СО2, которое вырабатывается в
организме за то же самое время.
Основные расчеты. Если заключение об
интенсивности энергетического обмена должно быть сделано на основании данных о
количестве поглощаемого кислорода, ход рассуждении состоит в следующем. Выход
энергии при окислении глюкозы описывается уравнением
С6Н12О6
+ 602 –> 6СО2+ 6Н2О + 2826 кДж. (2)
В данной реакции 2826 кДж–это полная энергия (энтальпия),
выделяемая при окислении 1 моль глюкозы; только часть этой энергии (свободная
энтальпия [5]) может быть использована для обеспечения клеточных функций.
Энергетическая ценность. Количество вырабатываемой
энергии часто выражают в соответствии с массой или объемом субстрата; масса 1
моля глюкозы равна 180 г, а объем 6 моль кислорода –6 х 22,4 л = 134,4 л. Из
этого следует, что полное окисление 1 г глюкозы сопровождается выделением 2826/180
= 15,7 кДж. Следовательно, энергетическая ценность глюкозы составляет
15,7 кДж/г .
Энергетический эквивалент («калорический эквивалент»)
выражает количество вырабатываемой энергии в соответствии с количеством
поглощенного кислорода. В случае приведенной выше реакции эта величина равна
2826 кДж/134,4 л = 21,0 кДж на 1 л O2.
Поскольку смесь углеводов, присутствующих в обычной пище, имеет несколько более
высокую энергетическую ценность по сравнению с глюкозой, энергетический
эквивалент окисления углеводов составляет 21,1 кДж на 1 л О2(табл.
24.4).
Дыхательный коэффициент (или коэффициент легочного
газообмена) указывает на тип пищевых продуктов, использованных в обмене
веществ; этот показатель определяется следующим образом:
ДК= VCO2/VO2 =Выделение
СO2/ Потребление O2 (3)
В случае окисления глюкозы потребление кислорода
равно выделению диоксида углерода, так что ДК = 1. Таким образом, значение
ДК, равный 1,–характерный показатель окисления углеводов.
Таблица 24.4. Дыхательный коэффициент (ДК)
и энергетический эквивалент при окислении питательных веществ (1 кДж
≈0,24 ккал)
|
|
Углеводы
|
Жиры
|
Белки
|
|
ДК
|
1,00
|
0,70
|
0,81
|
|
кДж/л O2
|
21,1
|
19,6
|
18,8
|
Пример расчета. В условиях покоя поглощение кислорода составляло
280 мл/мин (стандартный объем при стандартных значениях температуры, давления и
влажности воздуха), а значение ДК оказалось необычным–равным 1,00.
Интенсивность обмена веществ в этом случае равна 0,280·21,1 = 5,91 кДж/мин я: 8510
кДж/сут (≈98 Вт). Сходным образом определяют ДК в случае окисления жиров.
Поскольку в жирных кислотах на один атом углерода приходится меньше атомов
кислорода, чем в углеводах, их окисление характеризуется значительно более
низким дыхательным коэффициентом (0,7). В случае окисления чисто белковой
пищи ДК оказывается равным 0,81 (табл. 24.4).
Конечные продукты катаболизма. К конечным продуктам катаболизма наряду с другими
компонентами относятся вода (примерно 350 мл в день), диоксид углерода
(примерно 230 мл/мин), оксид углерода (примерно 0,007 мл/мин), мочевина (около
30 г/сут) и ряд азотсодержащих веществ (около 6 г/сут), а также другие
соединения, выделяющиеся с мочой.
Мочевина–это типичный конечный продукт распада белков,
поэтому по количеству мочевины и других азотсодержащих веществ можно оценивать
интенсивность катаболизма белков. При смешанной диете на белки в среднем
приходится 16% азота в пище, так что, умножив количество азота, присутствующего
в моче, на величину 6,25, можно оценить количество катаболизированного белка.
Уровень белкового обмена, необходимого
главным образом для поддержания структуры организма и его роста, поддерживается
примерно постоянным. При сбалансированном питании у жителей Центральной
Европы на белки обычно приходится примерно 15% всего энергетического
обеспечения. [Структурный белковый обмен существенно возрастает при несчастных
случаях и после операций, в связи с чем был введен термин «постагрессионный
метаболизм» (табл. 24.6).] Пропорции жиров и углеводов в пище, напротив,
значительно колеблются, так что различия в значениях дыхательного коэффициента
обусловлены в основном этими пищевыми компонентами. Итак, ДК можно
использовать для вычисления той части энергопродукции, которая связана с катаболизмом
жиров и углеводов, а также для определения того, сколько энергии
вырабатывается при поглощении 1 л кислорода (табл. 24.5). Изменение ДК на 0,1
соответствует изменению энергетического эквивалента на
Таблица 24.5. Зависимость энергетического
эквивалента от дыхательного коэффициента (ДК) без учета вклада белков (15%) в
общий метаболизм. Средний ДК составляет 0,82
|
ДК
|
1,0
|
0,9
|
0,82
|
0,8
|
0,7
|
|
кДж/л O2
|
21,1
|
20,6
|
20,2
|
20,1
|
19,6
|
0,5 кДж/л O2. Данные, приведенные в табл.
24.5, могут быть использованы для непрямого определения точных значений
интенсивности метаболизма.
Пример расчета. Поглощение кислорода, как и в предыдущем
примере, составляло 280 мл/мин, а значение ДК равнялось 0,82 (среднее значение
при величине энергетического эквивалента, равной 20,2 кДж/л O2). В
этом случае интенсивность метаболизма равна 0,280 • 20,2 ≈ 5,66 кДж/мин ≈ 8150 кДж/сут
(приблизительно 94 Вт). Разница между этим значением и результатом в предыдущем
примере составляет 358 кДж/сут, или 4%.
Факторы, влияющие на ДК. Соотношение
между количествами выделяемого углекислого газа и потребляемого кислорода
зависит от следующих трех факторов.
1. Тип питательных веществ, участвующих в обмене.
Как было указано, ДК равен: при окислении углеводов–1,0, при окислении
жиров–0,7 и при окислении белков–0,81 (табл. 24.4).
2. Гипервентиляция. Дополнительное количество
СO2, выдыхаемое при гипервентиляции, поступает из тех обширных
запасов СO2, которые содержатся в тканях и крови, и не связано с
образованием углекислоты в ходе метаболических процессов. Гипервентиляция не
влияет на объем поглощаемого кислорода, поскольку кровь и ткани организма не
могут накапливать дополнительный кислород. В переходной фазе,
предшествующей установлению нового, более низкого парциального давления СО2
в тканях и крови, ДК заметно возрастает и в некоторых случаях
достигает величины, равной 1,4. Гипервентиляцию могут вызывать такие факторы,
как произвольная активность (например, надувание воздушного баллона),
нереспираторный ацидоз (например, во время и после изнурительной работы),
психологический стресс (например, состояние предельного возбуждения) и
искусственно осуществляемое дыхание, при котором минутный объем вентиляции превышает
требуемый уровень.
3. Превращение одних питательных веществ в
другие. В тех случаях, когда большую часть рациона составляют углеводы,
последние могут превращаться в жиры. Поскольку в жирах кислорода содержится
меньше, чем в углеводах, этот процесс сопровождается выделением
соответствующего количества кислорода. Так, в случае перенасыщения углеводами
количество поглощаемого в легких кислорода снижается, а ДК возрастает.
В предельных случаях насильственного питания были
зарегистрированы такие значения дыхательного коэффициента, как 1,38 у гусей и
1,59 у свиней. В периоды голодания и при диабете ДК может падать
до 0,6. Это связано с усилением интенсивности обмена жиров и белков при
снижении метаболизма глюкозы (потребление запасов гликогена или нарушение утилизации).
При непрямом определении интенсивности обмена
веществ, если нет полной уверенности в соответствии между «респираторным
ДК» и условиями катаболизма («метаболический ДК»), следует использовать
среднее значение энергетического эквивалента, равное 20,2 кДж/л О2 и
соответствующее величине метаболического ДК 0,82. Как видно из табл. 24.5,
диапазон колебаний энергетического эквивалента в зависимости от значений ДК не
очень велик, поэтому погрешность, связанная с использованием среднего значения
энергетического эквивалента, не превышает ±4%.
Измерение
интенсивности обмена веществ в отдельных органах
Потребление кислорода (и, следовательно,
интенсивность метаболизма) в отдельных органах можно определить методом,
основанным на принципе Фика . В соответствии с этим принципом измеряют скорость
перфузии органа, v’O2, и артериовенозную разницу
по О2 и СO2:
v’O2 (мл/мин) =
Q’ (мл/мин) • (FaO2 – FbO2) (4)
v’CO2 (мл/мин) =
Q’ (мл/мин)
• (FвСO2 – Fa СO2)– (5)
В ткани головного мозга метаболизируются в основном углеводы,
и ДК примерно равен 1,0; ДК для скелетных мышц, а также для сердечной мышцы
существенно варьирует в зависимости от особенностей обмена веществ в данной
ситуации.
24.5. Измерение
интенсивности поглощения кислорода во всем организме
При определении интенсивности обмена веществ
непрямыми методами необходимо измерять поглощение кислорода испытуемым в
единицу времени. Для этой цели используют как закрытые, так и открытые
респираторные системы.
Закрытые системы
Принцип таких систем состоит в том, что испытуемый
вдыхает содержимое заполненного кислородом спирометра рис. 24.7).
Выдыхаемую газовую смесь пропускают через резервуар, в котором поглощается
диоксид углерода, после чего газовая смесь вновь поступает в спирометр;
таким образом,
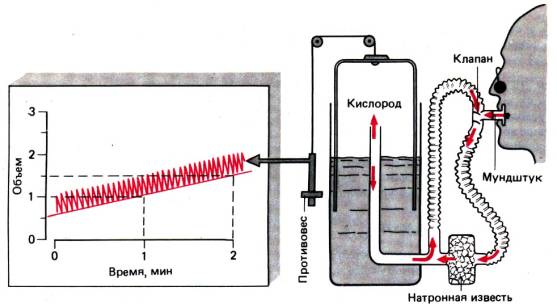
Рис. 24.7. Принцип закрытой системы для
измерения поглощения кислорода. Испытуемый вдыхает кислород из
колоколообразного газометра: СO2 поглощается натронной известью и
удаляется из выдыхаемого воздуха до возращения в газометр. Проведя линию через
нижние точки колебательной кривой и определив угол наклона этой линии, можно
найти значение скорости поглощения кислорода (в данном случае 0,5 л/мин)
путь газовой смеси оказывается замкнутым, а
респираторная система–закрытой. Регистрируемая спирограмма характеризуется углом наклона (рис.
24.7); чем круче наклон, тем больше кислорода удалено из системы в единицу
времени.
Закрытые системы должны быть заполнены
кислородом, поскольку при использовании воздуха кислород расходуется так
быстро, что его концентрация во вдыхаемой газовой смеси быстро падает,
становясь ниже 8,5 мл/дл (критический порог), при этом концентрация СО2 не
увеличивается. Нехватка кислорода в этих условиях часто вызывает неожиданную
потерю сознания у испытуемого, из–за того что дыхание стимулируется лишь в незначительной
степени, а другие сигнальные системы обычно не срабатывают . Преимущество
закрытых систем состоит в том, что нет необходимости измерять концентрацию O2,
а недостаток–в том, что нельзя определить ДК.
Стандартный объем. Какими бы ни были условия, в которых измерена
интенсивность поглощения кислорода, результаты нужно пересчитать на стандартные
условия, чтобы можно было сравнивать данные, полученные в разных экспериментах,
независимо от действительных значений температуры и давления воздуха. Принимаются
следующие условия СТДВ (стандартные температура, давление, влажность):
0°С, 760 мм рт.ст. и сухой воздух. Коэффициент пересчета обычно получают на
основе табличных данных по уравнению
Vo=V· Pб–PH2O
/760·273/273+t (6)
где Vo– объем в стандартных
условиях, V измеряемый объем, Pб– барометрическое давление,
РH20
давление водяного пара в спирометре, t–температура измеряемого
объема газа в градусах Цельсия.
Открытые системы
В открытых респираторных системах пути, по которым
следует вдыхаемый и выдыхаемый воздух, разделены. Обычно вдыхается свежий
воздух, а на пути выдыхаемого воздуха установлен прибор для измерения его объема,
а также концентрации в нем О2 и СO2. Поскольку
содержание соответствующих компонентов во вдыхаемом воздухе известно, можно
вычислить потребление кислорода и выделение углекислого газа.
Основные расчеты. При измерении в нормальных условиях можно
вполне удовлетворительно оценить поглощение О2 и выделение СО2 следующим
образом (см. [8]):
v’O2 = v’e·( FIO2–FEO2) (7)
или
v’СO2 = v’e· FEСO2) (8)
В том случае, если требуется особенно высокая
точность, следует иметь в виду, что истинные значения потребления О2 и
выделения СО2 выражаются разностью концентраций каждого из этих
веществ во вдыхаемом и выдыхаемом воздухе. Содержание СО2 в свежем
воздухе в нормальных условиях так мало, что им можно пренебречь. Но выдыхаемый
воздух содержит значительное количество O2, и для оценки потребления
О2 по разнице его концентраций во вдыхаемом и выдыхаемом воздухе
может быть использована следующая формула:
v’O2 = v’i FIO2– v’·FEO2) (9)
Количество выдыхаемого кислорода можно вычислить так
же, как и количество выдыхаемого СO2. Содержание О2 во вдыхаемом воздухе, когда используется свежий воздух,
известно (FiO2 = 0,2095 = 20,95 мл/дл), но V’I
неизвестно. Только если ДК = 1, можно считать, что минутные объемы
вдыхаемого и выдыхаемого воздуха (приведенные к стандартным условиям)
одинаковы. Когда ДК ниже 1,0, выдыхается меньше воздуха, чем вдыхается. Однако,
когда известен ДК, минутные объемы можно определить один из другого. Эти
соображения позволяют рассчитать потребление кислорода при использовании камеры
Дугласа (см. ниже) по следующей формуле (вывод уравнения не приводится):
DV’O2 = V’E(1,265 • DFO2– 0,265 • FEСO2) (10)
Величину v’e можно
определить по количеству выдыхаемого воздуха, собранного в камере Дугласа за
определенное время; FEСO2 –это фракция СО2 в собранном смешанном выдыхаемом воздухе, а DFO2
представляет собой разность между фракциями кислорода во вдыхаемом воздухе и
выдыхаемой газовой смеси. После этого результат следует пересчитать на
стандартные условия СТДВ.
Камера Дугласа. Камера Дугласа [11] применяется в одном из
классических методов для определения количества потребляемого кислорода. Метод
заключается в непрерывном измерении с помощью портативного
устройства, которое можно подсоединять к испытуемому, не стесняя его поведения.
При физиологических исследованиях испытуемый может носить камеру в процессе
работы как рюкзак. Свежий воздух поступает через клапан, расположенный в
мундштуке; при вдохе нос зажимается специальным зажимом. Весь
выдыхаемый воздух собирается в газонепроницаемой камере, куда он
поступает по системе клапанов и трубок; при этом время заполнения камеры точно
регистрируется. После окончания периода заполнения с помощью специальных
манипуляций добиваются тщательного перемешивания компонентов поступившего в
камеру воздуха, а затем определяют в пробе содержание кислорода и диоксида
углерода. Объем всего собранного воздуха измеряют с помощью газометра.
Другие методы. Вместо камеры Дугласа на спине у испытуемого можно
закреплять газометр, специально приспособленный для того, чтобы пробы
выдыхаемой газовой смеси попадали в маленькую дополнительную емкость. Эти пробы
должны быть репрезентативными в отношении всех компонентов (например,
мертвого пространства и альвеолярного воздуха) выдыхаемой смеси (так называемая
кратная порция). Поскольку в период выдоха пропорции различных компонентов
могут изменяться, необходимо, чтобы определенная часть воздушного потока
постоянно направлялась в отводную емкость (рис. 24.8); таким образом собирают
определенную часть выдыхаемой газовой смеси, в которой составляющие компоненты
представлены в адекватных соотношениях (например, 1% в портативной системе
Мюллера и Франса [15]).
Для непрерывного измерения поглощения
кислорода применяют более сложные методы и стационарные установки. Можно также
использовать современную телеметрическую аппаратуру на подвижной платформе.
Установки для непрерывной регистрации
поглощения кислорода обычно построены на основе одного из двух принципов,
проиллюстрированных на рис. 24.8. Прибор, в котором применен принцип кратной
порции,– пневмотахограф–(рис. 24.8,АI) используют обычно для анализа
непрерывного потока выдыхаемого воздуха. Однако концентрации газов определяются
не путем непрерывного измерения химической абсорбции, а при помощи газового
анализатора, действие которого основано на использовании определенных
физических свойств кислорода и диоксида углерода. Метаболические параметры
рассчитывают при этом так же, как и в методе с применением камеры Дугласа.
Главные преимущества установок, основанных на
принципе постоянного отсоса (рис. 24.8, Б), заключаются, во–первых, в том, что
можно просто и очень точно измерить при помощи газометра однородные воздушные
потоки, и, во–вторых, в том, что изменения концентраций О2 и СО2 в выдыхаемом воздухе
непосредственно отражают изменения исследуемых метаболических параметров. Таким
образом, наблюдаемые изменения концентраций газов прямо характеризуют
метаболический статус в данных конкретных условиях, так что не требуется
сложных вычислений, увеличивающих возможность ошибки.
При непрерывной регистрации важно, чтобы разность
концентраций газов и соответствующие объемы измерялись точно в один и тот же
момент времени. Поэтому необходимо как можно чаще проверять эту аппаратуру;
для калибровки удобен в качестве метода сравнения
метод с применением камеры Дугласа. В некоторых случаях следует использовать
специальные приемы, имитирующие потребление кислорода,–удаление кислорода при
помощи регулируемого пламени [16] и разбавление азотом [17].
Если нужно измерить интенсивность обмена веществ у
больного в условиях искусственного дыхания, необходимо учитывать возможность
изменений содержания кислорода во вдыхаемом воздухе, а в некоторых случаях и
добавление наркотических газообразных веществ [17].
24.6. Диагностическое значение показателей энергетического обмена
Физиологические и клинические
аспекты физической нагрузки. Физическое напряжение,
испытываемое человеком при работе или занятиях спортом, во многих случаях можно
выразить с помощью показателей энергетического обмена. Поскольку известны
средние значения соответствующих показателей (см. обзоры [4] и [6]), многие
виды работы можно классифицировать по затрачиваемым усилиям, как показано на
рис. 24.4.
Клиническая диагностика. В условиях шока (при
критическом падении артериального давления, например, в результате сильной
кровопотери) интенсивность обменных процессов падает ниже уровня
Клапан
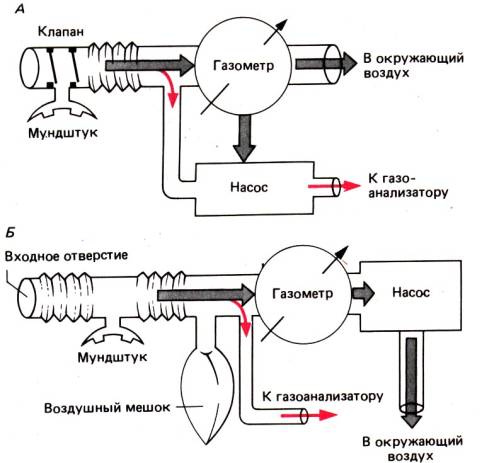
Рис. 24.8. Принципы непрерывного
измерения поглощения кислорода и выделения углекислого газа в открытых системах.
А. Принцип кратной порции. Испытуемый вдыхает свежий воздух через
мундштук с двусторонним клапаном; нос при этом зажат специальным зажимом.
Количество выдыхаемого воздуха измеряют с помощью газометра (или
пневмотахографа), который регулирует всасывающий насос таким образом, что
небольшая порция выдыхаемого воздуха, по величине пропорциональная скорости
газового потока, отводится и собирается. Это устройство позволяет получать репрезентативные
газовые пробы, содержащие пропорциональные количества всех компонентов
выдыхаемого воздуха («кратная порция»). 6. Принцип постоянного отсоса.
Испытуемый вдыхает и выдыхает газовую смесь через мундштук без клапанов (либо
через маску или капюшон), в то время как насос постоянно прогоняет у мундштука
поток свежего воздуха (в количестве, большем, чем это требуется для дыхания). У
ответвления, идущего к газовому анализатору, поток смешанного воздуха
однороден, поскольку зависит только от интенсивности работы насоса, а не от
минутного дыхательного объема. Когда быстро выдыхается большой объем воздуха,
он может временно сохраняться в воздушном баллоне. Поскольку выдыхаемый воздух
не выходит из системы через входное отверстие и работа насоса равномерна,
непрерывно измеряемые изменения концентраций кислорода и углекислого газа у
ответвления, идущего к газовому анализатору, пропорциональны поглощению
кислорода и выделению углекислого газа. Расчеты см. в [8]
основного обмена из–за недостаточности кровотока в
периферических областях. Возникает кислородный долг; во многих клетках
организма уровень метаболизма перестает обеспечивать их нормальное функциональное
состояние. По мере усиления периферического кровотока при ослаблении шока
интенсивность обменных процессов возрастает; таким образом, по изменению
интенсивности обмена веществ можно судить о тяжести шокового состояния.
Интенсивность обмена веществ во время болезни.
В случаях травм, ожогов и при высокой температуре
интенсивность обмена существенно изменяется (табл. 24.6). В связи с
постагрессионным метаболизмом она увеличивается, при этом в результате усиления
белкового обмена количество азота, выделяемого с мочой, возрастает в четыре
раза или более. Подобные изменения следует учитывать при назначении больным
искусственных питательных смесей.
Нарушения функции щитовидной железы также отражаются на уровне
обмена веществ: если ее активность чрезмерно высока (гипертиреоз),
интенсивность основного обмена повышается, а если активность щитовидной железы
чрезмерно низка (гипотиреоз), интенсивность обмена снижается (описание
соответствующей клинической картины. В отдельных случаях интенсивность обмена
веществ может отклоняться более чем на +100 или —40% от нормального уровня.
Однако в настоящее время показатели интенсивности основного обмена для
диагностики заболеваний щитовидной железы больше не используются.
Таблица 24.6. Увеличение интенсивности
обменных процессов (по отношению к обмену веществ в покое) и выделения
азотистых соединений с мочой (100%=
0,085 г/кг •сут) у разных больных [14]
|
Категории больных
|
Возрастание%
|
|
интенсивность обмена
|
выделение
|
|
1. Хирургические
операции средней тяжести
|
24
|
150
|
|
2. Множественные
переломы с другими повреждениями
(дорожные происшествия)
|
32
|
275
|
|
3. Огнестрельные
ранения
|
37
|
280
|
|
4. То же, что в п. 2,
отягощенное повреждением головы, требующим стероидной терапии
|
61
|
300
|
|
5. Заражение крови
|
79
|
330
|
|
6. Ожоги с большой
площадью поражения
|
132
|
335
|
24.7. Литература
Учебники
и руководства
1. Consolazio
C.F., Johnson R.E., Pecora L.J. Physiological measurements of metabolic
functions in man. New York–Toronto–London. McGraw–Hill, 1963.
2. Kestner
0., Knipping H. W. Die Ernahrung des Menschen. Berlin. Springer, 1924.
3. Lavousier
A. L., de LaPlace P. S. Abhandlung iiber die Warme (Erstveroffentlictiung
1780). In: Rosenthal J. (ed.). Zwei Abhandlungen uber Warme. Leipzig.
Wilhelm Engelmann, 1892.
4. Lehmann
G. Energetik des arbeitenden Menschen. In: Lehmann G. (ed.).
Handbuch der gesamten Arbeitsmedizin, Bd. 1. Arbeitsphysiologie.
Berlin–Munchen–Wien. Urban & Schwarzenberg, 1961.
5. Opitz
E., Lubbers D. Allgemeine Physiologic der Zeil– und Gewebsatmung. in: Buchner
F., Letterer E., Roulet F. (eds.). Handbuch der allgemeinen Pathologie, Bd–
4, Teil II. Der StofTwechsel. Berlin–Gottingen–Heidelberg. Springer, 1957.
6. Spitzer
H., Hettinger Th., Kaminsky G. Tafein fur den Energieumsatz (6. vollst.
uberarb. AufL). Berlin–Koln. Beuth., 1982.
7. Stegemann
J. Leistungsphysiologie –
Physiologische Grundlagen der Arbeit und des Sports. Stuttgart. Thieme, 1984.
8. Ulmer
H.–V. Zur Methodik, Stantardisierung und Auswertung von Tests fur die
Prufung der korperlichen Leistungs–fahigkeiten. Koln. Deutscher Artzeverlag,
1975.
9.
Wissenschaftliche Tabellen – Documenta Geigy (ed. J.R. Geigy A.G., Pharma, Basel). 7. Аufl. Basel, 1969.
Оригинальные статьи и обзоры
10. Boothby
W.M., Berkson J., Dunn H.L. Studies of the energy of metabolism of normal
individuals. A standard of basal metabolism, with a nomogram for clinical
application. Amer J. Physiol, 116, 468 (1936).
11. Douglas
C.G. A method for determining the total respiratory exchange in man. J.
Physiol. (London), 42, 17(1911).
12. Gopfert
H., Bernsmeier A.. Slufler R. Uber die Steigerung des EnergiestofTwechsels
und der Muskelinnervation bei geistigcr Arbeit. Pfliigers Arch., 256, 304
(1953).
13. Harris
J. A.. Benedict F. G. A biometric study of basal metabolism in man. Publ.
Nr. 279. Carneg. Inst., Washington 1919, cited in Stegemann,
Leistungsphysiologie. Stuttgart. Thieme, 1971.
14. Long
С. L., Schaffel N., Geiger J. W., Schiller W. R.,
Blakemore W.S. Metabolic response to injury and illness. Estimation of energy
and protein needs from indirect calorimetry and nitrogen balance. J. Parent.
Ent. Nutr., 3, 452 (1979).
15. Muller
E.A., Franz H. Energieverbrauchsmessungen bei beruflicher Arbeit mit einer
verbesserten Respirationsgasuhr. Arbeitsphysiologie, 14, 499 (1952).
16. Stegemann
J., Essfeld D. Advantages of the computerized breath–by–breath method for the
interpretation of spiroergometric data. In: Lollgen H,, Mellerowicz H.
(eds.). Progress in ergometry. Quality control and test criteria, p. 30,
Berlin–Heidelberg–New York–Tokyo. Springer, 1984.
17. Semsroth
M. Indirckte Kalorimetrie bei beatmeten Kindern. 2. Teil. Ein Messverfahren
und seine Uberprufung an einem neuentwickelten Stoffwechsel–Lungenmodell.
Infusionstherapie, 12, 294 (1985).