ТКАНЕВОЕ ДЫХАНИЕ
Й. Гроте
23.1.
Тканевой метаболизм и потребности тканей в кислороде
Обмен
веществ и преобразование энергии в клетках.
Тканевым дыханием называют обмен
дыхательных газов, происходящий в массе клеток при биологическом окислении
питательных веществ. В ходе окислительных процессов клетки поглощают из
капилляров кислород и одновременно выделяют конечный продукт метаболизма
– углекислый газ [8]. Здесь термин «тканевое дыхание» использован в более
широком смысле, чем в большинстве пособий по биохимии, где им обозначают
окислительный распад питательных веществ с участием молекулярного кислорода.
Поскольку недостаток O2 лимитирует окислительные реакции значительно сильнее, чем их
лимитирует неадекватное удаление СO2, мы будем рассматривать прежде всего процессы,
обеспечивающие снабжение тканей кислородом.
Аэробные и анаэробные процессы. Каждая живая клетка нашего организма нуждается в
поступлении определенного количества энергии. Эта энергия необходима для
поддержания нормальной структуры и жизнедеятельности клеток, а также для
выполнения специфических функций. В нормальных условиях клетки получают энергию
главным образом путем окислительного (аэробного) разложения питательных
веществ. Для осуществления аэробного метаболизма в клетке должны поддерживаться
определенные концентрации субстратов (углеводов, белков и жиров) и молекулярного
кислорода.
В анаэробных условиях необходимая клетке энергия
может быть получена только в процессе гликолиза. Конечный продукт гликолиза–лактат–заключает
в себе еще много энергии, поэтому данный путь метаболизма менее экономичен, чем
аэробное разложение глюкозы. Для получения одного и того же количества энергии
в анаэробных условиях в клетке должно расщепляться примерно в 15 раз больше
глюкозы, чем в аэробных. Согласно Бертону и Кребсу [15], при окислительном
распаде 1 моль глюкозы в условиях, примерно соответствующих внутриклеточной
среде (t = 25 °С, рН 7,0, PO2 = 159 мм рт. ст. = 20 кПа, PCO2 = 50 мм рт. ст. = 5,3 кПа),
высвобождается около 689 ккал = 2883 кДж свободной энергии. При разложении того
же количества глюкозы в процессе гликолиза выделяется лишь 50 ккал = 208
кДж свободной энергии. Несмотря на низкий энергетический выход, анаэробный
распад глюкозы играет важную роль как в анаэробных, так и в аэробных условиях
во многих тканях (например, в мозговом веществе почек, хрящах, клетках
сетчатки, эритроцитах и работающих мышцах). При изучении обмена веществ в
работающих скелетных мышцах, а также во внутренней зоне и сосочках мозгового
вещества почек было показано, что в противоположность существовавшим ранее
представлениям гликолиз может протекать с высокой скоростью даже в аэробных
условиях [17, 18].
Биологическое
окисление в митохондриях
Биологическое окисление происходит в митохондриях.
Кроме ферментов цикла лимонной кислоты, дыхательной цепи и окислительного
фосфорилирования в этих органеллах были обнаружены также ферменты расщепления
жирных кислот и ряда аминокислот [4]. На рис. 23.1 схематично показаны
различные пути окислительного метаболизма в митохондриях.
Пируват, жирные кислоты и аминокислоты
переносятся из цитоплазмы через митохондриальные мембраны в матрикс
митохондрий. После ряда биохимических превращений все эти субстраты
распадаются до веществ, которые поступают в цикл лимонной кислоты.
Пируват, образующийся главным образом в процессе аэробного расщепления глюкозы
в цитоплазме, превращается в результате окислительного декарбоксилирования в
матриксе митохондрий в ацетилкофермент А (ацетил–КоА), большая часть которого в
нормальных условиях распадается в цикле лимонной кислоты. В отличие от
расщепления глюкозы, первый этап которого (гликолиз) протекает в цитоплазме,
окислительный распад жирных кислот полностью происходит в матриксе митохондрий.
Молекулы
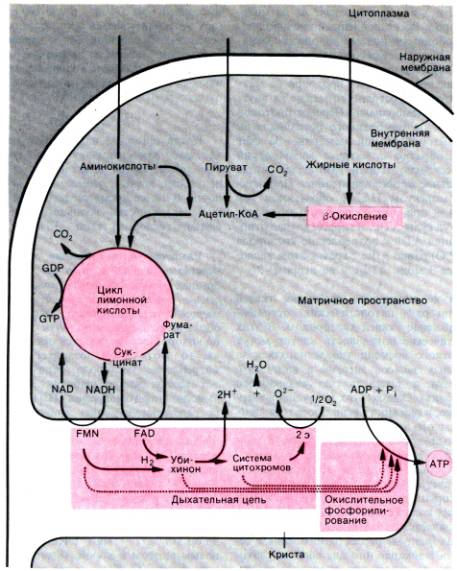
Рис. 23.1. Схема важнейших этапов
биологического окисления в митохондриях
жирных кислот окисляются в цепи биологических
реакций, называемой b–окислением; образующийся при
этом ацетил–КоА поступает в цикл лимонной кислоты либо используется для синтеза
жирных кислот. Органические кислоты, образовавшиеся в матриксе митохондрий при
метаболизме аминокислот, могут включаться в цикл лимонной кислоты на разных
этапах в виде ацетил–КоА, a–кетоглутарата, сукцинил–КоА,
фумарата и оксалоацетата.
Образовавшиеся в цикле
лимонной кислоты молекулы НАДН (NADH, продукт трех реакций
дегидрогенизации) и сукцинат диффундируют из матрикса митохондрий к их
внутренней мембране, в которой локализованы ферменты дыхательной цепи и
окислительного фосфорилирования. Здесь НАДН подвергается окислению комплексом ферментов
дыхательной цепи, включающим в качестве кофермента флавинмононуклеотид (FMN).
Окисление сукцината осуществляется комплексом ферментов дыхательной цепи, в
котором роль кофермента выполняет флавинадениндинуклеотид (ФАД, FAD).
Оба флавопротеиновых комплекса передают электроны на убихинон (кофермент Q),
который затем окисляется с участием цитохромов. В конечном счете из каждой
молекулы водорода образуется 2 протона и 2 электрона. Далее электроны
переносятся специфическими железосодержащими компонентами дыхательной цепи
(цитохром b, FeS–белок, цитохром С1, цитохром С2) на
цитохромоксидазу (комплекс цитохромов A и A3), которая передает их на
молекулярный кислород. Восстановленный кислород соединяется со свободными
ионами водорода с образованием воды. Энергия, высвобождающаяся в процессе
переноса электронов по дыхательной цепи, используется для перекачивания
протонов из матрикса митохондрий в пространство между их внутренней и наружной
мембранами. Перенос протонов приводит к созданию мембранного потенциала на
внутренней митохондриальной мембране. За счет тока протонов обратно в матрикс
митохондрий индуцируется, согласно теории Митчелла, синтез АТФ (окислительное
фосфорилирование). При окислении каждой молекулы НАДН образуются три молекулы
АТФ, а при окислении одной молекулы ФАДН2–две молекулы АТФ. В первом
случае отношение образования АТФ к потреблению кислорода (коэффициент Р/0)
равно 3 (на 1 грамм–атом кислорода образуются 3 моль АТФ), а во втором–2.
Последствия недостаточного снабжения тканей
кислородом.
При ряде патологических состояний нарушается снабжение тканей кислородом. В
этих случаях энергетические потребности клеток могут в течение короткого
времени удовлетворяться за счет ограниченных запасов энергии в виде АТФ и
креатинфосфата, а также за счет анаэробного процесса–гликолиза. Однако этих
источников энергии недостаточно, и они могут использоваться лишь в течение
небольшого периода времени, что вызвано двумя основными причинами. Во–первых, в
анаэробных условиях потребность клеток в глюкозе настолько
возрастает, что чаще всего не может полностью удовлетворяться в течение
длительного времени. Во–вторых, в процессе гликолиза образуется в больших
количествах лактат, который не столь быстро удаляется из ткани для
использования в других органах (например, для расщепления в печени, почках или
миокарде или для синтеза гликогена). При сильном недостатке кислорода
содержание лактата в тканях и крови постоянно возрастает, что в конечном счете
приводит к нереспираторному ацидозу. Когда рН внутриклеточной среды
падает ниже уровня, оптимального для активности ферментных систем, наступают
резкие нарушения в клеточном метаболизме.
Потребность тканей в кислороде
Потребление O2 в состоянии покоя. Количество кислорода,
потребляемого тканью, зависит от функционального состояния входящих в ее состав
клеток. В табл. 23.1 приведены данные о потреблении кислорода различными
органами и их частями, когда организм находится в состоянии покоя при
нормальной температуре. Скорость потребления кислорода тем или иным органом (V’O2 ) обычно выражают в мл O2 на 1 г или 100 г массы за 1
мин (при этом учитывается масса органа в естественных условиях). В соответствии
с принципом Фика V’O2
определяют,
исходя из кровотока (Q) через тот или иной орган и
разницы в концентрациях O2 в
поступающей к органу артериальной крови и оттекающей от него венозной крови
(авРO2 ):
V’O2=авРO2·Q’ (1)
Когда организм находится в
состоянии покоя, кислород относительно интенсивно поглощается миокардом, серым
веществом головного мозга (в частности, корой), печенью и корковым
веществом почек. В то же время скелетные мышцы, селезенка и белое вещество
головного мозга потребляют меньше кислорода (табл. 23.1).
Различия в потреблении кислорода разными участками
одного и того же органа. Во многих органах можно измерить кроваток через
ограниченные участки ткани путем определения клиренса инертных газов
(например, 85Кr, 133Хе и Н2).
Таким образом, если возможно взять пробу крови из вены, по которой
осуществляется отток от данного участка, то этот метод позволяет определить в
нем потребление кислорода. Кроме того, несколько лет назад был разработан метод
позитронной эмиссионной томографии (ПЭТ), позволяющий непосредственно измерять
кровоток и потребление O2 в отдельных частях органов. Этот метод успешно применяется для
исследования головного мозга человека [21, 36]. До внедрения метода ПЭТ, как
видно из табл. 23.1, измерить региональное потребление O2 можно было лишь в немногих
органах.
При изучении потребления кислорода тканями мозга
различных млекопитающих было показано, что кора больших полушарий потребляет от
8•10–2 до 0,1 мл O2 • г –1 • мин –1.
Исходя из потребления O2 целым мозгом и корой, можно вычислить
среднее потребление O2 белым веществом мозга. Эта величина составляет примерно 10–2
мл –1· мин–1. Прямое измерение поглощения O2 участками головного мозга у
здоровых испытуемых методом позитронной эмиссионной томографии дало следующие
величины: для серого вещества (в различных участках)–примерно от 4 до 6·
10–2 мл –1· мин–1, для белого
вещества–2·10–2 мл –1·
мин–1 [21, 36]. Можно предполагать, что потребление кислорода варьирует
не только в зависимости от участка, но также в разных клетках одного участка. В
самом деле, при измерении (с помощью платиновых микроэлектродов) регионального
потребления O2 поверхностными клеточными
слоями коры головного мозга было показано, что в условиях слабого наркоза это
потребление в пределах небольших участков варьирует примерно от 4–10–2
мл до 0,12 мл–г–1 •мин–1.
Таблица 23.1. Средние значения скорости
кровотока (U), артериовенозной разницы по O2 (авРO2)
и потребления
O2 (VO2 ) в различных органах
человека при 37 °С
|
Орган |
Q’, мл г-1 мин-1 |
авРO2 |
V’O2, мл г-1 мин-1 |
Источник данных |
|
|
Кровь |
- |
- |
0,6 ·10-4 1·10-4 |
[22] |
|
|
Скелетные
мышцы: |
|||||
|
в покое |
2·10-2-4·10-2 |
10·10-2—15·
10-2 |
2,5·10-3-5·10-3 |
[3, 32] |
|
|
при
тяжелой физической
нагрузке |
0,5-1,3 |
|
0,1-0,2 |
|
|
Селезенка:
|
1,0 |
1-10-2 |
1-10-2 |
[8, 42] |
|
|
Головной мозг: |
0,4-0,6 |
6·10-2-7·10-2 |
3·10-2-4·10-2 |
[2, 4, 19, 21, 27, 36] |
|
|
кора |
0,6-1,0 |
10·10-2 |
5·10-2-10·10-2 |
||
|
белое вещество |
0,2-0,3 |
5·10-2-7·10-2 |
1·10-2-2·10-2 |
||
|
Печень: |
1,0 (25%-по печеночной артерии) |
4·10-2-5·10-2 (воротная
вена- печеночная
вена) 8·10-2-10·10-2 (печеночная
артерия- печеночная вена) |
5·10-2-б·10-2 |
[8, 24, 35] |
|
|
Почки: |
4,0 |
1,5·10-2-2·10-2 |
5,5·10-2-6,5·10-2 |
[10, 29] |
|
|
корковое
вещество |
4,0-5,0 |
2·10-2-2,5·10-2 |
9·10-2-10·10-2 |
||
|
наружный
слой мозгового
вещества |
1,2 |
5-10-2 |
6·10-2-6,5·10-2 |
||
|
внутренний слой мозгового
вещества |
0,25 |
1·10-2-2·10-2 |
0,3·10-2-0,5·10-2 |
||
|
Сердце: |
[1, 7, 13, 20, 26, 40] |
||||
|
в
покое |
0,8-0,9 |
10·10-2-15·10-2 |
7·10-2-10·10-2 |
||
|
при
тяжелой физической нагрузке |
до
4,0 |
До
≈17·10-2 |
До
≈ 40·10-2 |
||
Результаты радиоавтографических исследований
регионального кровотока (с использованием иод–14C–антипирина)
и регионального потребления глюкозы (с использованием 14C–2 –дезоксиглюкозы)
в коре головного мозга позволяют считать, что эти параметры также существенно
различаются в соседних участках [37, 38]. У людей старше 30 лет региональный
кровоток и потребление O2 в сером веществе головного мозга с возрастом постепенно снижаются [19,
21, 36]. Примерно такие же различия в потреблении кислорода были обнаружены
между отдельными частями почек. В корковом веществе почек среднее
потребление O2 в несколько раз больше, чем
во внутренних участках и сосочках мозгового вещества. Поскольку
потребности почек в кислороде зависят главным образом от интенсивности активной
реабсорбции Na+ из просвета канальцев в ткани, полагают, что столь
выраженные различия в региональном потреблении O2 обусловлены в основном разницей между
величинами этой реабсорбции в корковом и мозговом веществе [11].
Потребление O2 в условиях повышенной
активности органа. В
том случае, если активность какого–либо органа по тем или иным причинам
повышается, в нем возрастает и скорость энергетического обмена, а
следовательно, и потребность клеток в кислороде. При физической нагрузке
потребление O2 тканями миокарда
может увеличиваться в 3–4 раза, а работающими скелетными мышцами–более чем
в 20–50 раз по сравнению с уровнем покоя. Потребление O2 тканями почек
возрастает при увеличении скорости реабсорбции Na+.
В большинстве органов скорость поглощения O2 не зависит от скорости
кровотока в них (при условии, что напряжение O2 в тканях достаточно велико). Почки
составляют исключение. Существует критическая скорость перфузии, превышение
которой вызывает образование ультрафильтрата; при этом уровне фильтрации
повышенный кровоток сопровождается повышенным потреблением O2 тканью почек. Такая
особенность обусловлена тем, что интенсивность клубочковой фильтрации (а
следовательно, и реабсорбции Na+ пропорциональна скорости
кровотока.
Зависимость потребления О2 от температуры. Потребление O2 тканями крайне
чувствительно к изменениям температуры. При снижении температуры тела
энергетический обмен замедляется, а потребность большей части органов в
кислороде уменьшается. При нормальной терморегуляции активность органов,
участвующих в поддержании теплового баланса, увеличивается, а потребление ими
кислорода возрастает. К таким органам относятся, в частности, скелетные мышцы;
их терморегуляторная функция осуществляется за счет повышения мышечного тонуса
и дрожи. Увеличение температуры тела сопровождается повышением потребности большинства
органов в кислороде. Согласно правилу Вант–Гоффа, при изменении температуры на
10°С в пределах от 20 до 40 °С потребление тканями кислорода изменяется в том
же направлении в 2–3 раза (Q10 = 2–3). При некоторых
хирургических операциях бывает необходимо временно остановить кровообращение (а
следовательно, снабжение органов O2 и питательными
веществами). При этом, чтобы уменьшить потребности органов в кислороде, часто
используют гипотермию (понижение температуры тела): больному дают такой
глубокий наркоз, при котором терморегуляторные механизмы подавляются.
23.2. Снабжение тканей
кислородом
Запасы кислорода в тканях
Количество кислорода, которое
клетки могут использовать для окислительных процессов, зависит от величины конвекционного
переноса O2 кровью и уровня диффузии
O2 из капилляров в ткани.
Поскольку единственным запасом кислорода в большинстве тканей служит его
физически растворенная фракция, снижение поступления O2 с кровью приводит к тому,
что потребности тканей в кислороде перестают удовлетворяться, развивается
кислородное голодание и замедляются обменные процессы.
Миоглобин как депо
кислорода.
Единственная ткань, в которой имеются более или менее существенные запасы
кислорода,–это мышечная. Роль депо кислорода в мышечной ткани выполняет
пигмент миоглобин (Mb), способный обратимо
связывать O2. Однако содержание
миоглобина в мышцах человека невелико, поэтому количество запасенного кислорода
не может обеспечить их нормального функционирования в течение длительного
периода кислородного голодания.
На примере миокарда особенно
хорошо видно, насколько ограничены запасы O2, связанного с
миоглобином. Среднее содержание миоглобина в сердце составляет 4 мг на 1 г
ткани. Поскольку 1 г миоглобина может максимально связать 1,34 мл кислорода, в
физиологических условиях запасы кислорода в мышце сердца составляют около
0,5–10–2 мл O2 на 1г ткани. Этого количества в условиях
полного прекращения снабжения O2 может хватить лишь для того, чтобы
поддерживать окислительные процессы в течение примерно 34с.
Значение миоглобина в снабжении мышечной ткани
кислородом. Миоглобин играет роль кратковременного депо и внутриклеточного
переносчика O2. Обратимо
связывая O2, он служит своего рода
кислородным буфером. Благодаря такому буферному эффекту различия в
парциальном давлении O2 в
разных участках мышц выражены гораздо меньше, чем в тканях, не содержащих
миоглобин, а в
условиях нагрузки, несмотря на увеличение
потребления O2, его парциальное давление
поддерживается почти постоянным. Сродство миоглобина к кислороду по сравнению
со сродством гемоглобина очень велико; поэтому миоглобин выполняет функцию
кислородного буфера лишь при парциальных давлениях O2, меньших чем 10–15 мм
рт.ст. (1,3–2,0 кПа). При тех условиях, которые создаются в скелетных мышцах,
парциальное давление полунасыщения O2 для миоглобина составляет около 5–6 мм рт.ст.
(0,7–0,8 кПа). Благодаря высокому сродству миоглобина к O2 в мышечных клетках
поддерживается низкое парциальное давление этого газа, что приводит к созданию
значительного градиента парциального давления O2 между мышечными клетками и капиллярной
кровью, способствующего высвобождению O2 в мышцах [22, 25]. Функция миоглобина как
переносчика O2 проявляется тогда, когда в
мышечной клетке возникают градиенты концентрации кислорода. Молекулы
оксигенированного миоглобина диффундируют из областей с высоким содержанием O2 в области с его низким
содержанием (облегченная диффузия O2). При этом небольшие градиенты парциального
давления O2 могут вызывать
внутриклеточный перенос больших количеств этого газа [22, 30].
В миокарде кислород, связанный с миоглобином, обеспечивает
протекание окислительных процессов в тех участках, кровоснабжение которых на
короткий срок снижается или полностью прекращается во время систолы [26].
В начальном периоде интенсивной мышечной нагрузки
возросшая потребность скелетных мышц в O2 частично удовлетворяется за счет кислорода,
высвобождаемого миоглобином. В дальнейшем возрастает мышечный кровоток, и
поступление кислорода к мышцам вновь становится адекватным. Количество
кислорода, освобожденного оксимиоглобином, составляет часть кислородного
долга, который должен быть восполнен в каждом мышечном волокне.
Поступление кислорода к тканям и его утилизация
Поступление кислорода к
органам.
Количество O2, доставляемое кровью к тому
или иному органу за единицу времени, равно произведению концентрации O2 в артериальной крови и
скорости кровотока:
Поступление
O2 = CaO2 · Q. (2)
Из этого уравнения видно, что
различия в поступлении кислорода к органам связаны только с различиями в их
кровоснабжении. Любое изменение скорости кровотока, возникающее в результате
изменений периферического сопротивления или среднего артериального давления,
приводит непосредственно к изменениям в снабжении тканей кислородом.
Среднюю величину поступления O2 к тому или иному органу в
нормальных условиях можно вычислить, исходя из содержания O2 в артериальной крови и
величины кровотока в этом органе (см. табл. 23.1). Особенно обильно снабжаются
кислородом корковое вещество почек, селезенка и серое вещество головного мозга.
Поступление O2 к скелетным мышцам в
состоянии покоя, мозговому веществу почек и белому веществу головного мозга
невелико.
Коэффициент утилизации O2 в разных органах.
Коэффициент утилизации O2 тем или иным органом равен
отношению потребления O2 к его поступлению. Этот коэффициент можно вычислить на основе
уравнений (1) и (2):
коэффициент
утилизации O2 =авРСO2·Q’/CaO2·Q’ = авРСO2/CaO2 (3)
Этот коэффициент варьирует в
соответствии с различными потребностями тканей в кислороде. При нормальных
условиях кора головного мозга, миокард и скелетные мышцы в состоянии покоя
потребляют около 40–60% поступающего кислорода. В норме коэффициент утилизации O2 для организма в целом
составляет 0,3. В условиях повышенной активности коэффициент утилизации O2 может существенно
возрастать. Так, при крайне интенсивной физической нагрузке коэффициент
утилизации O2 работающими скелетными
мышцами и миокардом достигает 0,9. При патологических состояниях утилизация O2 тем или иным органом может
значительно увеличиваться в результате снижения содержания O2 в артериальной крови (артериальной
гипоксемии) или уменьшения кровоснабжения органа (ишемии). Особенно мал
коэффициент утилизации O2 в почках и селезенке. Это связано с тем, что потребности данных
органов в кислороде невелики, а кровоток (и, следовательно, поступление O2) в них значителен в связи с
их особыми функциями.
Обмен
дыхательных газов в тканях
Свободная и облегченная
диффузия.
Обмен дыхательных газов между капиллярной кровью и тканями происходит, как и в
легких, путем диффузии. Молекулы O2 движутся по градиенту напряжения (парциального
давления) этого газа из эритроцитов и плазмы в окружающие ткани. Одновременно
происходит диффузия углекислого газа, образующегося в окислительных процессах,
из тканей в кровь (напряжение СO2 в тканях велико, а в крови
мало). Энергией, обеспечивающей диффузию дыхательных
газов, служит кинетическая энергия отдельных молекул этих газов. Таким образом,
напряжение O2 и СO2 в крови играет большую роль в
тканевом газообмене. Средние значения напряжений этих газов в различных частях
кровеносного русла человека в условиях покоя представлены на рис. 23.2.
На высвобождение O2 из крови в ткани может
влиять диффузия оксигемоглобина внутри эритроцитов, благодаря которой
ускоряется перенос молекул O2 к поверхности этих клеток [30]. Этот процесс называется облегченной
диффузией кислорода. В мышцах аналогичным образом влияет на перенос O2 диффузия оксимиоглобина.
При определенных условиях – очень низком парциальном давлении O2 и соответственно
незначительными градиентами этого давления внутри клеток – облегченная диффузия
кислорода играет существенную роль в кислородном обеспечении мышечных клеток
[22].
Переносу дыхательных газов
может также способствовать конвекция цитоплазмы в эритроцитах или
клетках ткани, а также плазмы и интерстициальной жидкости.
На перенос дыхательных газов
(O2–из крови в ткани, СO2–в обратном направлении)
влияют не только градиенты их напряжения между кровью и клетками, но
также площадь поверхности обмена, величины диффузионного расстояния
и диффузионного сопротивления тех сред, через которые осуществляется
перенос. Зависимость диффузионного потока того или иного газа от всех этих
факторов при постоянном градиенте напряжения (или концентрации) описывается
первым законом диффузии Фика.
Модели газообмена в тканях. Закон диффузии можно использовать
для теоретического анализа обмена дыхательных газов в тканях и вычисления их
напряжений в клетках. Подобные построения основаны на математических моделях,
включающих различные параметры, от которых зависит диффузия в участках ткани,
снабжаемых одним капилляром или капиллярной сетью. Для того чтобы можно было
описать процессы диффузии O2 и СO2 математически,
функциональные и морфологические условия, в которых она происходит, должны быть
заданы в упрощенном виде.
Для анализа газообмена в тканях созданы различные
структурные модели. Наиболее широко известна и чаще всего используется модель
тканевого цилиндра, предложенная в 1918 г. Крогом [31]. В этой
модели участок ткани, снабжаемый одним капилляром, рассматривается как цилиндр,
осью которого служит этот капилляр. Крог исходил из данной модели в своих
исследованиях диффузии O2 в скелетных мышцах. Несмотря на то что модель Крога может описывать
условия обмена газов достаточно точно лишь для того случая, когда капилляры
участка ткани лежат параллельно друг другу, начинаются и заканчиваются в одной
плоскости и несут кровь в одном и том же направлении, она оказалась чрезвычайно
полезной для изучения обмена дыхательных газов и других веществ в тканях. В настоящее
время имеется и ряд других моделей, в том числе так называемая конусная
модель, основанная на допущении, что соседние капилляры несут кровь в
противоположных направлениях. Предложена также кубическая модель, в которой
рассматривается участок ткани между четырьмя параллельными капиллярами с
противоположным направлением кровотока; другие модели описывают процесс
газообмена в слое ткани, содержащем капиллярную сеть с квадратными
ячейками.
Зависимость газообмена от
плотности капилляров и особенностей микроциркуляции. На обмен дыхательных газов
в участке ткани влияет не только градиент напряжения этих газов между
капиллярной кровью и клетками, но также плотность капилляров и распределение
кровотока в микроциркуляторном русле. От числа перфузируемых капилляров, их
длины и расстояния между ними непосредственно зависят как площадь
диффузионной поверхности, на которой происходит газообмен между кровью
и тканью, так и диффузионное расстояние
внутри ткани.
Плотность капилляров варьирует не только в
зависимости от органа, но иногда даже в пределах одного и того же органа. В
тканях, характеризующихся высоким уровнем метаболизма, имеется плотная
капиллярная сеть, что способствует газообмену.
В миокарде, например, на каждое мышечное волокно приходится
один капилляр; среднее расстояние между соседними капиллярами составляет около
25 мкм. В коре головного мозга это расстояние равно примерно 40 мкм, а в
скелетных мышцах – приблизительно 80 мкм. Распределение кровотока в
микроциркуляторном русле зависит от величины тонуса гладких мышц сосудов,
предшествующих капиллярам. Изменение тонуса этих сосудов влияет на количество
одновременно перфузируемых капилляров. Таким образом, от площади диффузионной
поверхности и величины диффузионного расстояния зависит не только поступление кислорода
к какому–либо участку ткани, но также условия обмена O2.
Напряжение (парциальное
давление) O2 в тканях
Критическое напряжение O2 в митохондриях.

Рис. 23.2. Напряжение O2 (РO2)
и СO2 (РCO2) в крови в различных участках сосудистой
системы в условиях покоя (по [41] с изменениями)
Напряжение O2 в клетках в норме имеет промежуточное значение между
напряжением O2 в артериальной крови и
минимальным значением, которое в органах (или частях органов) с высокими
потребностями в кислороде составляет около 1 мм рт.ст. (133,3 Па). Для
нормального протекания окислительных процессов необходимо, чтобы напряжение O2 в области митохондрий
превосходило 0,1–1 мм рт.ст. (13,3–133,3 Па). Эта величина называется
критическим напряжением O2 в митохондриях [15, 22, 39]. Если напряжение O2 в участках цитоплазмы, непосредственно граничащих с
митохондриями, падает ниже критического, то полное окисление восстановленной цитохромоксидазы
становится невозможным, перенос водорода и электронов в дыхательной цепи
подавляется и в результате не может поддерживаться нормальная скорость
энергетического обмена. Таким образом, важнейшим показателем,
характеризующим снабжение тканей кислородом, служит напряжение O2 в клетках.

Рис. 23.3. Напряжение O2 в
сером веществе коры головного мозга. А. Гистограмма регионального
напряжения O2 в коре головного мозга морской свинки в условиях
нормальной вентиляции легких; максимальное напряжение O2 в
исследуемой ткани такое же, как и в артериальной крови PO2= 90–95 мм рт. ст.
=12,0–12,7кПа); минимальное PO2 (в тех клетках, которые хуже
всего снабжаются кислородом) составляет 0,5 1 мм рт. ст. (67–133 Па), что
примерно на 25 мм рт. ст. (3,3 кПа) ниже среднего напряжения O2 в
венозной крови сосудов коры головного мозга [34]. Б. Устройство
микроэлектрода для полярографического измерения напряжения O2 в
тканях
Внедрение в практику
исследований полярографических методов позволило непосредственно
измерять напряжение O2 в отдельных клетках при помощи микроэлектродов. Напряжение O2 в клетках, расположенных
близко к поверхности, определяют при помощи миниатюрных платиновых
микроэлектродов, введение которых в ткань не приводит к нарушению в ней
микроциркуляции. Для измерения напряжения O2 в более глубинно расположенных клетках используют
игловидные электроды с диаметром кончика в пределах 0,5–5 мкм (рис. 23.3, Б).
Оба этих метода используют в
основном в опытах на животных. В то же время они были успешно применены и при
исследовании больных для определения напряжения O2 в легкодоступных органах. Так, подробно
изучено распределение напряжения O2 в покое и при нагрузке в пораженных мышцах при ряде
мышечных заболеваний и состояний, сопровождающихся нарушением мышечного
кровотока. При нейрохирургических операциях с помощью поверхностных микроэлектродов
получены важные данные, касающиеся поступления кислорода к тем или иным
участкам головного мозга. Результаты подобного исследования представлены на
рис. 23.4 в виде гистограмм парциального давления O2 в поверхностных клетках
различных участков коры головного мозга в условиях артериальной нормоксии и
артериальной гипоксии [28].
В большинстве случаев,
однако, снабжение кислородом какого–либо органа у человека рассчитывают по
результатам непосредственного измерения важнейших показателей, влияющих на
поступление O2,–скорости кровотока,
напряжения и концентрации дыхательных газов, рН артериальной крови; на
основании этих данных анализируют газоообмен в интересующем участке ткани.
Распределение напряжения O2 в ткани мозга. Наибольший интерес представляет
распределение напряжения O2 в ткани головного мозга и в миокарде, поскольку при недостаточном
поступлении кислорода к любому из этих двух органов может наступить смерть.
Среднее распределение напряжения O2 в цилиндрическом участке коры головного мозга, снабжаемом
одним капилляром, представлено на рис. 23.5 (при этом потребление O2 принимается равным 9–10–2 мл·г·мин–1, а
кровоток–0,8 мл·г·мин–1). При
прохождении крови через капилляр
напряжение O2 в нем падает с 90 мм рт.
ст. (12,0 кПа) примерно до 28 мм рт. cm. (3,7 кПа). Эти
изменения соответствуют эффективной кривой диссоциации оксигемоглобина.
Перпендикулярно продольному градиенту напряжения направлен радиальный градиент
напряжения с разницей между напряжением O2 в крови и в периферических участках цилиндра около
26 мм рт. ст. (3,5 кПа). Хуже всего снабжаются кислородом клетки, расположенные
у венозного конца цилиндра;

Рис. 23.4. Гистограммы регионального
парциального давления O2 в клетках, расположенных у поверхности коры
головного мозга кошки в условиях артериальной нормоксии (А; РaO2
= 96 мм рт. ст.= 12,8 кПа), умеренной артериальной гипоксии (Б; РaO2 = 52 мм рт. ст. = 7,0 кПа) и
тяжелой артериальной гипоксии (В;PaO2 =31 мм рт. ст.= 4,2 кПа). По
мере снижения РO2 в артериальной крови распределения сдвигаются в
сторону все более низких величин РO2, что проявляется в значительном
увеличении числа измеренных значений PO2 в пределах 0–5 мм рт. ст.
(0–0,7 кПа). При тяжелой артериальной гипоксии возникает выраженная тканевая гипоксия
с аноксией многих клеток коры головного мозга [28]
по расчетам напряжение O2 в области этих клеток
составляет 1–2мм рт.ст. (133–266 Па).
Вычисленные величины напряжения O2 хорошо согласуются с
данными прямых измерений у животных [27] в аналогичных условиях (рис. 23.3. A и
23.4) и свидетельствуют о том, что ткань мозга отнюдь не так хорошо снабжается
кислородом, как принято считать. Эти расчеты позволяют понять, почему
уменьшение мозгового кровотока столь легко приводит к кислородному голоданию
нейронов, расположенных в наиболее плохо снабжаемых кровью участках. В
результате функция таких нейронов быстро нарушается, что во многих случаях
приводит к частичной или полной потере сознания.
Распределение напряжения O2 в миокарде. Сердечная мышца отличается
от большинства других тканей тем, что снабжение ее кислородом носит
периодический характер. В ходе сердечного цикла изменяются как потребность
миокарда в энергии, так и его кровоснабжение. При систоле в результате
повышенного интрамурального давления кровоток в бассейне левой коронарной
артерии снижается и может на короткое время полностью прекратиться во
внутренних слоях миокарда левого желудочка. В результате снабжение миокарда
кислородом претерпевает периодические колебания: в систоле оно минимально,
а в диастоле максимально. В то же время потребность клеток миокарда в
энергии изменяется противоположным образом: она возрастает во время
фазы сокращения и снижается во время фазы расслабления.
Существуют два механизма, полностью удовлетворяющие
в нормальных условиях потребность миокарда в энергии, несмотря на снижение
поступления O2 во время систолы. Один из
них

Рис. 23.5. Схема распределения
напряжения O2 в модели тканевого цилиндра по Крогу; в качестве
примера приведен цилиндрический участок ткани коры головного мозга человека,
снабжаемый одним капилляром (потребление O2 =9•10–2мл•г–1•мин–1;
кровоток =0,8 мл•г–1•мин–1). В нормальных условиях
среднее напряжение O2 в крови понижается с 90 мм рт. ст. (12,0 кПа)
в области артериального конца капилляра до ~28 мм рт. ст. (3,7 кПа) в области
венозного конца. Имеется также радиальный градиент среднего напряжения O2,
направленный от капилляра к поверхности цилиндра и составляющий около 26 мм рт.
ст. (3,5 кПа)
заключается в том, что в тех клетках, где во время
сокращения РО2 падает ниже примерно 10 мм рт.ст. (1,3 кПа), миоглобин
играет роль кратковременного депо O2 (см. выше). Второй механизм состоит в том, что
временно повышенная потребность миокарда в энергии удовлетворяется за счет энергетических
резервов (АТФ и креатинфосфата). Во время систолы благодаря значительному
повышению кровоснабжения миокарда миоглобин снова полностью насыщается
кислородом, а клеточные запасы энергии восполняются [26].
Изменения в снабжении миокарда кислородом в ходе
сердечного цикла, по–видимому, сопровождаются периодическими колебаниями
напряжения O2 в клетках миокарда.
При физической нагрузке
создаются дополнительные трудности для нормального снабжения миокарда
кислородом. Сердце, выполняющее в этих условиях большую работу, потребляет и
большее количество O2. В то же время увеличение частоты сокращений
сердца приводит к значительному укорочению диастолы, и в результате нарушается
соответствие между снабжением сердца кислородом и потребностью в нем. В связи с
этим переносимость физической нагрузки ограничена предельной частотой
сокращений сердца, равной примерно 200 ударов в 1 мин. Действительно, на ЭКГ в
этих условиях часто появляются типичные симптомы гипоксии миокарда (снижение
сегмента ST, уплощение или инверсия зубца Т).

Рис. 23.6. Распределение парциального
давления O2 на поперечном срезе мышечного волокна, богатого
миоглобином и снабжаемого четырьмя капиллярами. Расчет сделан для следующих
условий физической нагрузки: потребление O2 0,14 мл•г–1•мин–1;
радиус волокна 25 мкм [25]
Распределение парциального
давления O2 в работающих скелетных
мышцах
Распределение парциального
давления O2 в работающих скелетных
мышцах в наибольшей степени зависит от функционирования миоглобина в
качестве буфера и переносчика O2. Теоретический анализ диффузии O2 в богатой миоглобином
мышечной ткани [25] показал, что при нагрузке могут создаваться очень большие
градиенты парциального давления O2 между кровью капилляров и снабжаемыми этими
капиллярами мышечными клетками (рис. 23.6). Когда парциальное давление O2 в мышечном волокне мало,
столь же невелики и градиенты PO2 в этом волокне;
следовательно, главным механизмом переноса O2 в мышечных клетках должна быть облегченная
диффузия. Результаты этих теоретических расчетов хорошо соответствуют значениям
внутриклеточного парциального давления, вычисленным на основе прямых измерений
насыщения миоглобина O2 в одиночных мышечных волокнах [22].
23.3 Регуляция снабжения
тканей кислородом и кислородное голодание
Механизмы, обеспечивающие
соответствие поступления кислорода потребности в нем
При усиленной деятельности
того или иного органа его повышенная потребность в O2 может удовлетворяться как
за счет увеличения снабжения кислородом, так и за счет более полной его
утилизации. Как следует из уравнения (2), поступление O2 к тканям может
увеличиваться за счет усиления кровотока либо повышения содержания O2 в артериальной крови или в
результате обоих этих эффектов. Однако повышение содержания O2 в артериальной крови путем
кратковременной гипервентиляции практически невозможно, поскольку в
физиологических условиях насыщение гемоглобина кислородом здесь уже составляет
около 97%. В связи с этим при временном увеличении потребности какого–либо
органа в кислороде доставка последнего возрастает главным образом за счет повышения
скорости кровотока.
Регуляция кровоснабжения органов. Количество крови,
протекающей через орган, зависит прежде всего от величины сердечного выброса
и тонуса сосудов, расположенных перед микроциркуляторным руслом этого
органа. Нервные, гуморальные и местные химические факторы, влияющие на
периферическое кровообращение, подробно рассмотрены в гл. 20. Здесь же мы
остановимся лишь на некоторых особенностях регуляции снабжения кислородом
мозга, сердца и скелетных мышц.
При усиленной деятельности мозга доставка к
нему O2 может возрастать главным
образом в результате снижения тонуса мышц сосудистых стенок. К факторам,
вызывающим расширение сосудов мозга, относятся снижение напряжения O2 (гипоксия) и повышение
напряжения СO2 (гиперкапния) во
внутриклеточном и внеклеточном пространствах, а также повышение концентрации
ионов Н+ во внеклеточном пространстве. Аналогичное
действие оказывает умеренное повышение содержания K+ во внеклеточном
пространстве и увеличение содержания аденозина в периваскулярном пространстве.
Влияние всех этих факторов уменьшается или полностью устраняется при понижении
содержания в периваскулярном пространстве ионов Са2+, так как эти
ионы играют центральную роль в обеспечении тонуса сосудов. Повышение концентрации
Са2 + во внеклеточной среде непосредственно вызывает
сосудосуживающий эффект, а ее снижение приводит к расширению сосудов [2, 4, 13,
33]. Остается неясным, какой вклад вносит в регуляцию мозгового кровотока
влияние вегетативных нервов на диаметр сосудов [2, 3].
При усиленной нагрузке на сердце кровоснабжение
миокарда увеличивается главным образом за счет влияния местных химических
процессов. Главным фактором их инициации служит снижение напряжения O2 в тканях (гипоксия).
Так, когда потребности сердца в кислороде превышают его поступление, в миокарде
увеличивается содержание аденозина вещества, оказывающего сосудорасширяющий
эффект. Возможно, именно аденозин играет основную роль в увеличении коронарного
кровотока. Кроме того, большое значение в местной регуляции кровоснабжения
миокарда играют эйкозаноиды. Особо важны в этом отношении простациклин
и простагландины, обладающие сосудорасширяющим действием [11]. Увеличение
тонуса симпатических нервов при нагрузке вначале приводит к снижению
кровоснабжения миокарда вследствие возбуждения α–адренорецепторов. Однако
одновременно активируются и b–адренорецепторы, что
вызывает усиление метаболизма и, как следствие, расширение сосудов. В итоге
увеличивается кровоснабжение миокарда (если перфузионное давление достаточно
велико) [1, 7, 20].
В вопросе о том, какие механизмы обеспечивают
повышение кровотока в скелетных мышцах, остается много неясного.
Известно, что эти мышцы иннервируются как адренергическими симпатическими
волокнами (от активности которых, как и во многих других органах, зависит тонус
гладких мышц сосудов), так и холинергическими симпатическими волокнами,
оказывающими сосудорасширяющее действие. Полагают, что первичное повышение
мышечного кровотока в начале физической работы связано именно с активацией
холинергических симпатических вазодилататоров. Влияние этих волокон на те
отделы микроциркуляторного русла, в которых происходит обмен дыхательных газов
и других веществ, выражено слабо. Кровоток в истинных (нутритивных) капиллярах
по мере продолжения работы мышц увеличивается, возможно, за счет действия
целого ряда местных химических факторов. Все они снижают базальный тонус
сосудистых мышц, не зависящий от нервных влияний. Полагают, что к таким
факторам относятся прежде всего повышение содержания К+ во
внеклеточной жидкости и увеличение ее осмотического давления. Кроме
того, дополнительное действие может оказывать гипоксия мышц. Что же
касается напряжения СO2 и содержания ионов Н+, то сосудорасширяющий эффект этих
факторов при работе мышц играет лишь второстепенную роль [3].
Последствия длительного или систематического
повышения потребности в кислороде. В том случае, если организм систематически испытывает
повышенную потребность в кислороде, сердце должно столь же систематически
усиливать свою деятельность. При этом в миокарде наступают структурные
изменения и масса сердца в целом увеличивается. Такие приспособительные
процессы наблюдаются, например, у спортсменов, занимающихся видами спорта,
требующими выносливости. Масса сердца у них может достигать 500 г (при норме
200–300 г). Это происходит главным образом за счет увеличения отдельных волокон
миокарда (гипертрофии). Полагают, что основным стимулятором роста
миокардиоцитов служит их кратковременное кислородное голодание.
Гипертрофия миокарда не может развиваться безгранично, так как при ней
ухудшается снабжение миокардиоцитов кислородом. В том случае, если масса сердца
превосходит критическое значение, равное примерно 500 г, увеличивается как
масса мышечной ткани, приходящаяся на один капилляр, так и количество
капилляров; однако прекапиллярные сосуды при этом изменяются незначительно. В
результате уровень поступления O2 к клеткам сердца снижается [б].
Предельный радиус миокардиоцита при гипертрофии составляет около 13,5 мкм (в
норме он равен примерно 8 мкм).
В патологических условиях масса сердца может
становиться больше критической. В результате наступает кислородное голодание
многих участков миокарда, разрушение части миокардиоцитов и нарушение
нормальной структуры сердца (патологическая гипертрофия сердца с
расширением, или дилатацией, его полостей). При систематическом повышении
потребности организма в кислороде могут наблюдаться не только изменения со
стороны сердца, но также увеличение кислородной емкости крови. В этих
условиях, как и при нехватке кислорода на больших высотах или нарушениях легочного газообмена,
происходит усиление эритропоэза и синтеза гемоглобина. Фактором,
запускающим эти процессы, служит увеличение продукции, главным образом в
почках, эритропоэтина – вещества, ускоряющего образование или созревание проэритробластов.
Возрастание числа эритроцитов (эритроцитоз) и концентрации гемоглобина приводит к повышению
кислородной емкости крови. При этом содержание O2 в артериальной
крови повышается, хотя его напряжение остается прежним. Однако вклад
эритропоэза в адаптацию очень ограничен, так как при повышении гематокрита
возрастает и вязкость крови, что вызывает дополнительную нагрузку на
сердце.
Причины
недостаточного снабжения тканей кислородом
При нарушениях газообмена в
легких или переноса дыхательных газов кровью может возникать состояние, при
котором доставка кислорода к тканям не соответствует их потребностям в O2. Такое состояние называют
тканевой гипоксией (если PO2 снижено) или тканевой
аноксией (если PO2= 0 мм рт. ст.). Среди возможных причин подобных
состояний основными являются следующие:
1) понижение напряжения O2 в артериальной крови (артериальная
гипоксия);
2) уменьшение кислородной
емкости крови (анемия);
3) уменьшение кровоснабжения
того или иного органа (ишемия).
Артериальная гипоксия. В том случае, если
вентиляционно–перфузионное соотношение в легких уменьшается (например, при
альвеолярной гиповентиляции, в артериальной крови падает как напряжение O2 (гипоксия), так и его концентрация (гипоксемия).
Одновременно повышается напряжение в артериальной крови СO2 (гиперкапния). и в результате развивается респираторный
ацидоз. Аналогичные сдвиги напряжения и концентрации O2 наблюдаются во время
пребывания на больших высотах, однако в этом случае напряжение СO2 в артериальной крови
понижается (гипокапния). и возникает респираторный алкалоз.
При тяжелой артериальной гипоксии снабжение тканей
кислородом ограничено, и в результате возможны лишь слабые физические нагрузки.
В этих условиях (и особенно в органах с высокими потребностями в O2) напряжение O2 в капиллярной крови может
резко уменьшаться; его падение приводит к венозной гипоксии (рис. 23.7 и
23.8). Из рис. 23.7 видно, что при выраженной артериальной гипоксии изменения
напряжения O2 в капиллярной крови в процессе
тканевого газообмена соответствуют участку с наиболее крутым наклоном на
эффективной кривой диссоциации оксигемоглобина. На этом участке кривой данному
изменению содержания O2 соответствует меньшее изменение напряжения O2, чем в области более пологого
правого участка кривой. В результате профиль напряжения O2 в капиллярах сильно
уплощен, хотя артериовенозная разница по кислороду та же, что и в норме.
Благодаря этому ухудшение снабжения тканей кислородом частично компенсируется.
На рис. 23.8

Рис. 23.7. Падение напряжения O2 в крови при
прохождении ее через капилляр в норме и при артериальной гипоксии (РO2 –40
мм рт. ст. 5,3 кПа). Данные приведены для миокарда (организм в состоянии
покоя). В том случае, если напряжение O2 в артериальной крови значительно
снижено, изменения этого напряжения в капиллярной крови соответствуют в
основном участку с крутым наклоном на кривой диссоциации оксигемоглобина.
Вследствие этого напряжение O2 уменьшается меньше, чем в условиях
нормоксии, что частично компенсирует неблагоприятные начальные условия
поступления O2 к тканям. По оси ординат–концентрация O2(СO2),
мл O2/мл крови; по оси абсцисс–напряжение O2 (РO2)
представлены кривые,
иллюстрирующие эти взаимосвязи на примере коры головного мозга. Напряжение O2 в артериальной крови в
данном случае принято равным 40 мм рт.ст. (5,3 кПа). Когда градиент напряжения O2 между кровью и тканями
становится слишком мал для того, чтобы кислород высвобождался в достаточном
количестве, внутриклеточное напряжение O2 в области венозного конца капилляра падает ниже
критического уровня РO2 в митохондриях, что приводит к угнетению
энергетического обмена.
Анемическая гипоксия. При кровопотере или
недостаточном синтезе гемоглобина (анемия), а также при образовании
в крови метгемоглобина или отравлении угарным газом (функциональная
анемия) уменьшается кислородная емкость крови. Это приводит к снижению
содержания в артериальной крови O2. Подобные нарушения иллюстрирует рис. 23.9, где в качестве
примера использован миокард. Если в данных условиях количество кислорода,
экстрагированного тканями, не изменяется, то содержание O2 в крови по мере прохождения
ее через капилляр существенно снижается. В области венозных концов капилляров
содержание O2 может достигать такого
низкого уровня, при котором

Рис. 23.8. Градиент среднего напряжения
O2 вдоль капилляра коры головного мозга человека в нормальных
условиях, при ишемической гипоксии (кровоток снижен на 1/3) и при тяжелой
артериальной гипоксии
наступает кислородное голодание тканей (венозная
гипоксия).
Ишемическая гипоксия. При нарушении
кровоснабжения какого–либо органа поглощение его клетками кислорода
увеличивается, и в результате возрастает артериовенозная разница по O2. Это приводит к увеличению
градиента напряжения O2

Рис. 23.9. Влияние анемии (Нb=10 г/дл) на изменения
напряжения O2 в капиллярной крови. Данные приведены для миокарда
(организм в состоянии покоя). По оси ординат–концентрация O2 (СO2),
мл O2/мл крови; по оси абсцисс–напряжение O2 (PO2)
вдоль капилляра (венозная гипоксия). Поскольку
одновременно снижается градиент напряжения O2 между кровью и тканями, снабжение клеток кислородом
может стать недостаточным (рис. 23.8).
Кислородотерапия;
кислородное отравление
Во многих случаях с
кислородным голоданием можно успешно бороться с помощью кислородотерапии. При
использовании этого метода стремятся повысить напряжение O2 в
артериальной крови путем увеличения парциального давления кислорода во
вдыхаемом воздухе. Для этого больному дают либо чистый кислород, либо смесь
газов с высоким содержанием O2 (изобарическая оксигенация): можно
также помещать больного в барокамеру, давление в которой поднимается выше
атмосферного (гипербарическая оксигенация). При ишемической или
анемической гипоксии кислородотерапия сравнительно малоэффективна, так как при
таких состояниях можно лишь незначительно повысить содержание O2 в
артериальной крови путем увеличения его физически растворенной фракции.
Кислородотерапию можно применять лишь в течение короткого
периода времени, поскольку длительное вдыхание газовой смеси с высоким
содержанием O2 приводит к кислородному отравлению. Так, при
нормальном барометрическом давлении чистым кислородом можно дышать не дольше 4
ч. Существенное повышение напряжения O2 в клетках (гипероксия)
сопровождается изменениями активности многих ферментов, участвующих в клеточном
метаболизме. Так, в условиях гипероксии угнетается окисление глюкозы, фруктозы
и пировиноградной кислоты. Классическими симптомами кислородного отравления
служат потеря сознания и судороги. При этом наблюдается также снижение
мозгового и почечного кровотока, а также уменьшение сердечного выброса
в связи с повышением тонуса блуждающих нервов. В легких происходят изменения
альвеолярной мембраны, что может сопровождаться нарушением процессов диффузии и
накоплением жидкости в интерстициальном пространстве и альвеолах (отек
легких). У новорожденных детей при длительном (в течении нескольких часов
или суток) лечении чистым кислородом наблюдаются поражения сетчатки,
приводящие к ухудшению зрения и даже полной слепоте.
Для того чтобы при длительной изобарической
оксигенации не наступило кислородное отравление, используют не чистый O2,
а газовую смесь. В смеси для взрослых содержание O2 не должно
превышать 0,6, а парциальное давление–величины 450 мм рт. ст. (60 кПа); в смеси
для новорожденных и грудных детей содержание O2 не должно превышать
0,4, а парциальное давление–300 мм рт. ст. (40 кПа) [10].
Обратимые
и необратимые нарушения при острой тканевой аноксии
Во всех случаях острой
тканевой аноксии–независимо от того, наступила она в результате внезапного
прекращения поступления O2 из–за закупорки сосуда или вследствие тяжелой артериальной
гипоксии–вслед за коротким латентным периодом, во время которого клетки
функционируют нормально, наступает период нарушения их метаболизма и функции.
По мере исчерпания запасов энергии клетки становятся неспособными поддерживать
даже самую минимальную активность, и в результате наступает полная утрата
функций. В то же время структура клеток, как видно из рис. 23.10, за счет
использования запасов энергии может сохраняться в течение нескольких минут и
даже часов (этот срок зависит от количества энергии, необходимой для
поддержания структур). Благодаря этому нарушения, происходящие в начальном
периоде аноксии, обратимы, и восстановление жизнедеятельности клеток
возможно. Когда энергетических запасов уже не хватает для сохранения клеточных
структур, развиваются необратимые нарушения, и в конечном . счете
наступает гибель клеток. В таких высокодифференцированных клетках, как нейроны,
необратимые изменения при температуре тела наблюдаются уже через 10 мин при
постоянной аноксии. В то же время в клетках скелетных мышц при таких же
условиях и нормальных запасах энергии необратимые изменения наступают лишь
через несколько часов.

Рис. 23.10. Изменения в тканевом метаболизме во время острой
ишемической аноксии и в восстановительном периоде. В нижней части рисунка
обозначены периоды времени после аноксии, в течение которых происходят
обратимые и необратимые изменения. Штриховая кривая соответствует динамике
восстановления нормальных обменных процессов после аноксии. В верхней части
рисунка обозначены характерные интервалы восстановительного периода
Утрата функций и их восстановление. Предел сохранения функций
(промежуток времени от начала тканевой аноксии до полной утраты функций), или
функциональный предел, короче по времени, чем предел оживления
(восстановления). Это означает, что после полной утраты функций какого–либо
органа в результате тканевой аноксии имеется период времени, в течение которого
возможно полное восстановление его жизнедеятельности.
Предел сохранения функций и предел оживления
особенно коротки в случае головного мозга. После полного прекращения
мозгового кровотока очень быстро–примерно через 4 с–возникают отдельные
нарушения функции мозга, а уже через 8–12 с (период сохранения функции)
наступает полная утрата его функций с потерей сознания. На ЭЭГ первые изменения
наблюдаются примерно через 4–6 с, а через 20–30 с спонтанная электрическая
активность головного мозга исчезает и регистрируется изоэлектрическая
электроэнцефалограмма.
При температуре 37 °С восстановить жизнедеятельность
головного мозга можно в течение 8–10 мин после наступления внезапной тканевой
аноксии–это предельный срок оживления головного мозга. Если аноксия была
очень кратковременной, то признаки функционирования мозга появляются примерно
через 1 мин после ее прекращения (латентный период восстановления). Если
же длительность ишемии мозга составляла около 4 мин, то латентный период
восстановления увеличивается приблизительно до 10 мин. Для полного же
восстановления функций часто необходимы часы или даже дни (период
восстановления). Даже в том случае, если продолжительность ишемии мозга
составляла лишь 1 мин, время восстановления достигает 15 мин. Предел оживления почек
и печени равен 3–4 ч, а период восстановления этих органов–нескольким
суткам. Восстановить жизнедеятельность сердца можно через несколько
часов, если оно не сокращалось, однако при активной деятельности уже через 3–4
мин после прекращения коронарного кровотока сердце теряет способность к
поддержанию гемодинамики на нормальном уровне. Если даже удается восстановить
деятельность сердца после внезапной остановки кровообращения, оно в течение 4–5
мин не может создать в артериальной системе давление, необходимое для
нормального мозгового кровотока, поэтому остановка сердца часто уже через
несколько минут приводит к необратимым изменениям в головном мозгу и к смерти.
В связи с этим предел реанимации всего организма составляет лишь около 4
мин, что значительно меньше, чем предел оживления отдельного жизненно важного
органа.
Учебники и руководства
1. Bassenge
E. Physiologic der Koronakdurchbluttung. In: Handbuch der inneren Medizin.
Bd. 9/3, S. 1. Berlin–Heidelberg–New York–Tokyo. Springer, 1984.
2. Betz E. Physiologic und Pathophysiologie der
Gehirndurchblutung. In: Handbuch der Medizinischen Radiologic, Bd. 14, S. 194,
Berlin, Heidelberg. Springer, 1981.
3. Golenhofen К. Skeletmuskel. In:
Lehrbuch der Physiologic in Einzeldarstellungen. Physiologic der Kreislaufs,
Bd. 1, S. 385. Berlin–Heidelberg–New York. Springer, 1971.
4. Hirsch H., Gehirn. In: Lehrbuch der Physiologic in
Einzeldarstellungen. Physiologic der Kreislaufs. Bd. 1, S. 145.
Berlin–Heidelberg–New York. Springer, 1971.
5. Lehninger A. I. Bioenergetik. Moleculare Grundlagen der
biologischen Energieumwandlungen. 2. Auflage, Stuttgart. Thieme, 1974.
6. Linzbach A.J. Pathologische Anatomic der Herzinsufii–zienz.
In: Handbuch der inneren Medizin., 4. Aufl., bd. 9, S. 706.
Berlin–Gottingen–Heidelberg. Springer, 1960.
7. Lochner W. Herz. In: Lehrbuch der Physiologic in
Einzeldarstellungen. Physiologic des Kreizlaufs. Bd. 1, S. 185,
Berlin–Heidelberg–New York. Springer, 1971.
8. Lutz J.. Bauereisen E. Abdominalorgane. In: Lehrbuch der
Physiologic in Einzeldarstellung. Physiologic der Kreizlaufs, Bd. 1, S. 229,
Berlin–Heidelberg–New York. Springer, 1971.
9. Piper J., Koepchen H. P. Atmung. In: Physiologic des Menschen.
Bd. 6. 2. Aufl. (Hrsg. Gayer 0. H., Kramer K., Jung R.). Munchen:
Urban u. Schwarzenberg, 1975.
10. Poulsen H., Jacobsen E. Die hyperbare Sauerstofftherapie. In:
Anaesthesiologie. Intensivmedizin und Reanimation. 5. Aufl. S. 805.
Berlin–Heidelberg–New York. Springer, 1982.
11. Schror К. Prostaglandine und
verwandte Verbindungen. Bildung, Funktion und pharmakologische Beeinflussung.
Stuttgart. Thieme, 1984.
12. Thurau К. Niere. In: Lehrbuch
der Physiologie in Einzeldarstellungen. Physiologie des Kreislaufs. Bd. 1, S.
293. Berlin–Heidelberg–New York. Springer, 1971.
Оригинальные статьи и обзоры
13. Betz
E. Cerebral blood flow: Its measurement and regulation. Physiol. Rev., 52,
595 (1972)
14. Bretschneider H. J. Sauerstoffbedarf und –versorgung des
Herzmuskels. Verh. dtsch. Ges. Kreisl.–Forsch., 27, 32 (1961).
15. Burton R., Krebs H.A. The free–energy changes associated with
the individual steps of the tricarboxylic acid cycle, glycolysis and alcohol
fermentation and with hydrolysis of the pyrophosphate groups of adenosintriphosphate.
Biochem. J., 54, 94 (1953).
16. Change В., Oshino N., Sugano Т., Mayevsky. Basic principles of tissue
oxygen determination from mitochondrial signals. Adv. Exp. Med. Biol., 37A, 277
(1973).
17. CohenJ.J. Is the function of the renal papilla coupled
exclusively to an anaerobic pattern of metabolism. Am. J. Physiol., 236, F423
(1979).
18. Connett R.J.. Gayeski Т.Е.]., Honig C.R. Energy sources in fully
aerobic rest–work transitions: a new role for glycolysis. Am. J. Physiol., 248,
H922 (1985).
19. Devous M. D. Sr., Stockely E. M„ Chehabi H. H., Bonte F. J. Normal
distribution of regional cerebral blood flow measurement by dynamic
single–photon emission tomography. J. Cereb. Blood Flow Metabol., 6, 95 (1986).
20. Feigl E. 0. Coronary Physiology, Physiol. Rev., 63, 1 (1983).
Physiol., 2, 274 (1967).
21. Frackowiak S.J.,
LenziG.L., Jones Т., Heather J.D. Quantitative measurement of regional cerebral blood flow and oxygen
metabolism in man using 15O and positron emission tomography:
theory, procedure and normal values. J. Comput. Tomogr., 4, 272 (1980).
22. Gayeski T.E.J., Connet R.J., Honig C.R. Oxygen transport in
restwork transition illustrates new functions for myoglobin. Am. J. Physiol.,
248, H914 (1985).
23. Greenbaum R., NunnJ.F., Pry s–Roberts D., KelmanG.R. Metabolic
changes in whole human blood (in vitro) at 37 °C. Respir. Physiol., 2, 274
(1967).
24. Greenway С. V., Stark R. D.
Hepatic vascular bed. Physiol. Rev., 51, 23 (1971).
25. Groebe K., Thews G. Theoretical analysis of oxygen supply to
contracted skeletal muscle. Adv. Exp. Med. Biol., 200, 495 (1986).
26. Grate J., Thews G. Die Bedingungen fur die
Sauerstoff–versorgung des Herzmuskelgewebes. Pflugers Arch., 276, 142 (1962).
27. Grole J., Zimmer К., Schubert R. Effects of severe arterial hypocapnia on regional blood flow
regulation, tissue PO;
and metabolism in the brain cortex of cats. Pflugers Arch., 391, 195
(1981).
28. Grote J., Schubert R. Regulation of cerebral perfusion and
PO; in normal and edematous brain tissue. In: Oxygen Transport to Human Tissue
(Eds. Loeppky J. A., Riedesel Af.L.), S. 169. New York, Amsterdam,
Oxford. Elsevier North Holland, 1982.
29. Kramer К., Thurau К.,
Deetjen P. Hamodynamie des Nierenmarks, 1. Mitteilung: Capillare
Passagezeit, Blut–volumen. Durchblutung, Gewebshamatokrit und O2–Ver–brauch
des Nierenmarks in situ. Pflugers Arch., 270, 251 (1960).
30. Kreuzer F. Facilitated diffusion of oxygen and its possible
significance: a review. Respir. Physiol., 9, 1 (1970).
31. Krogh A. The number and distribution of capillaries in
muscles with calculations of the oxygen pressure head nesessary for supplying
the tissue. J. Physiol. (Lond.), 52, 409 (1918/19).
32. Ките К. Das Sauerstoftdruckfeld im normalen und
pathologisch veranderten Muskel. In: Schriftenreihe Neurologie. Bd. 3.
Berlin–Heidelberg– New York. Springer, 1969.
33. Kuschinsky W., Wahl M. Local chemical and neurogenic
regulation of cerebral vascular resistance. Physiol. Rev., 58, 656 (1978).
34. Lubbers D. W. Local tissue PO2: its measurement
and meaning. In: Oxygen Supply. Theoretical and Practical Aspects of Oxygen
Supply and Microcirculation of Tissue (Eds. Kessler M., Bruley D. F., dark
L. C„ Lubbers D. W., Silver I. A., Sirmuss J.). S. 151. Munchen–Berlin–Wien.
Urban u. Schwarzenberg, 1973.
35. Lutz J., Henrich H„ Bauereisen E. Oxygen supply and uptake in
the liver and the intestine: Pflugers Arch., 360, 7 (1975).
36. Phelps M. E., Mazziotta J. С.,
Huang S.–C. Study of cerebral function with positron computed
tomography. J. Cereb. Blood Plow Metabol., 2, 113 (1982).
37. Sakurada 0., Kennedy С., Jehle J., Brown
J.D., Carbin G., SokoloffL. Measurement of local
cerebral blood flow with iodo [14C] antipyrine. Am. J. Physiol,,
234, H59 (1978).
38. SokoloffL., Reivich M., Kennedy C„ Des Rosiers M.H., Patlak C.
S., Pettrigrew K. D., Sakurada 0., Shinohara M. The [14C]
deoxyglucose method for the measurement of local cerebral glucose utilization:
theory, procedure, and normal values in the conscious and anesthetized albino
rat. J. Neurochem., 28, 897 (1977).
39. Starlinger H., Lubbers D. W. Polarographic measurements of
the oxygen pressure performed simultaneously with optical measurements of the
redox state of the respiratory chain in suspensions of mitochondria under
steady–state conditions at low oxygen tension. Pflugers Arch., 341, 15 (1973).
40. Strauer В. E.
Dynamik, Koronardurchblutung und Sauer–stoffverbrauch des normalen und kranken
Herzens. Experimentell–pharmakologische Untersuchungen und Kathete–runtersuchungen
am Patienten. Basel, Miinchen, Paris, London, New York, Sydney. S. Karger,
1975.
41. Thews G. Der Transport der Atemgase. Klin. Wschr., 41, 120
(1963).
42. Vaupel P., Wendling P., Thome H., Fischer J. Atemgas–wechsel
und Glucoseaufnahme der menschlichen Milz in situ. Klin. Wschr., 55, 239 (1977).