ФИЗИОЛОГИЯ ЧУВСТВА РАВНОВЕСИЯ, СЛУХА И РЕЧИ
Р. Клинке
Данная глава посвящена физиологии двух филогенетически
родственных сенсорных органов–слуха и
равновесия. Они не только тесно связаны анатомически, располагаясь рядом в
каменистой кости и образуя внутреннее
ухо, но и произошли в ходе эволюции из одной структуры. Поскольку важнейшее
для человека средство общения–речь–опосредовано органом слуха, физиология речи также рассмотрена в
данной главе.
Речь требует наличия слуха. Кроме того, словесное
общение–важнейшее средство обучения, поэтому глухота или даже лишь недостаток
слуха представляют собой наиболее серьезную угрозу для умственного развития
ребенка. Сравнительно–физиологические исследования показали, что глухота
сказывается на нем сильнее, чем слепота. Следовательно, слух–важнейшее для
человека чувство.
12.1. Физиология чувства равновесия
Физиология периферического сенсорного
аппарата
Вводные
анатомические комментарии.
Вестибулярный орган – одна из составных частей перепончатого лабиринта, образующего внутреннее ухо; другая его
составляющая–орган слуха (рис. 12.1). Перепончатый лабиринт заполнен жидкостью, эндолимфой, и погружен в другую,
называемую перилимфой. Вестибулярный
орган состоит из двух морфологических субъединиц – отолитового аппарата (macula utriculi и macula sacculi)
и полукружных каналов (передний и задний вертикальные и горизонтальный
каналы). В области макул (пятен) и в полукружных каналах вблизи от ампул
расположен содержащий рецепторы сенсорный эпителий, который покрыт желеобразной
массой, образованной в основном мукополисахаридами.
В отолитовом аппарате эта масса как бы подушкой покрывает сенсорные клетки и
содержит отложения карбоната кальция в форме крошечных кристаллов кальцита (отолитов). Благодаря наличию этих
«каменистых» включений она носит название
отолитовой мембраны. Буквальный перевод греческого термина "otolithus"–«yшнoй камень».
В полукружных каналах желеобразная масса больше напоминает мембранную
перегородку. Эта структура, купула, кристаллов
не содержит.
Рецепторы и адекватный стимул. В сенсорном эпителии макул и полукружных каналов
находятся два морфологически различных типа рецепторных клеток [8], которые,
очевидно, существенно не различаются своими физиологическими свойствами.
Оба типа клеток несут на свободной поверхности
субмикроскопические волоски (реснички),
поэтому называются волосковыми (рис. 12.2). С помощью электронного микроскопа
можно различить стереоцилии (по
60–80 на каждой рецепторной клетке) и
киноцилии (по одной). Рецепторы это вторичные сенсорные клетки, т. е. они
не несут собственных нервных отростков, а иннервируются афферентными волокнами
нейронов вестибулярного ганглия, образующими
вестибулярный нерв. На рецепторных клетках оканчиваются также эфферентные
волокна. Афференты передают в ЦНС информацию об уровне возбуждения рецепторов,
а эфференты изменяют чувствительность последних, однако значение этого влияния
до сих пор не совсем ясно [37]. Регистрация активности одиночных афферентных
волокон вестибулярного нерва показала их

Рис. 12.1.
Схема вестибулярного лабиринта. Его лимфатические пространства сообщаются с
улитковыми

Рис. 12.2.
Схема двух рецепторных клеток сенсорного эпителия вестибулярного органа и их
нервных волокон. При наклоне пучка ресничек в сторону киноцилии частота
импульсации в афферентном нервном волокне повышается, а при наклоне в
противоположную сторону снижается
относительно высокую регулярную активность покоя, т.е. импульсацию и в отсутствие внешних
стимулов. Если желеобразную массу экспериментально сдвигать относительно
сенсорного эпителия, такая активность увеличивается или уменьшается в
зависимости от направления смещения. Эти изменения происходят следующим
образом. Поскольку реснички погружены в желеобразную массу, при движении
последней они отклоняются. Сдвиг их пучка и служит адекватным стимулом для
рецептора. Когда он направлен в сторону киноцилии (рис. 12.2), активируется
соответствующее афферентное волокно: скорость его импульсации возрастает. При
сдвиге в противоположном направлении частота импульсов снижается [8]. Сдвиг в
направлении, перпендикулярном данной оси, активности не изменяет. Информация
передается из рецепторной клетки в окончание афферентного нерва за счет
рецепторного потенциала и неидентифицированного пока нейромедиатора [38].
Наиболее существенно здесь то, что сдвиг
(изгибание) ресничек–это адекватный стимул для вестибулярных рецепторов,
увеличивающий или уменьшающий (в зависимости от своего направления) активность афферентного
нерва. Таким образом, наблюдается морфологическая (по расположению ресничек) и
функциональная
(по характеру воздействия на активность) ориентация рецепторной клетки.
Естественные стимулы для макул. Как уже говорилось, реснички рецепторных клеток
погружены в отолитовую мембрану. У последней за счет присутствия кристаллов
кальцита плотность (приблизительно 2,2) существенно выше, чем у эндолимфы
(около 1), заполняющей остальную внутреннюю полость саккулуса (сферического
мешочка) и утрикулуса (эллиптического мешочка, маточки). Значит, вследствие
повсеместно присутствующего гравитационного ускорения всякий раз, когда
сенсорный эпителий отолитового аппарата не занимает совершенно горизонтального
положения, сила тяжести вызывает
скольжение (на очень малое расстояние) по нему всей отолитовой мембраны.
(Представьте себе, что произойдет, если желеобразная масса, обозначенная на
рис. 12.2 красным, очень тяжела, а вы, держа учебник вертикально, наклоняете
его вбок. Естественно, она будет соскальзывать под углом вниз.) Это перемещение
изгибает реснички, т.е. на рецепторы действует адекватный стимул [8]. Когда
человек стоит вертикально, а его голова находится в «нормальном» положении,
макула утрикулуса расположена почти горизонтально и отолитовая мембрана не прикладывает
сдвигового усилия к покрытому ею сенсорному эпителию. При наклоне головы макула
утрикулуса оказывается под углом к горизонту, ее реснички изгибаются и
рецепторы стимулируются. В зависимости от направления наклона частота
импульсации эфферентного нерва либо увеличивается, либо снижается. Ситуация с
макулой саккулуса в принципе аналогична, но она при нормальном положении головы
расположена почти вертикально (рис. 12.1). Таким образом, при любой ориентации
черепа каждая из отолитовых мембран по–своему воздействует на сенсорный
эпителий и возникает специфическая картина возбуждения нервных волокон.
Поскольку в каждой макуле две популяции рецепторных клеток с противоположно
ориентированными ресничками, нельзя сказать, что наклон головы в данном направлении
активирует афференты [8, 26]. Напротив, в любом случае одни волокна
активируются, а другие тормозятся. Такого положения головы, при котором
активность всех нервных волокон упала бы до нуля, не существует.
Центральные компоненты вестибулярной системы, оценивая
тип возбуждения вестибулярного нерва, информируют организм об ориентации черепа
в пространстве [3, 21, 26]. Обеспечение таких сведений–важнейшая функция
отолитовых органов. Гравитационное ускорение–лишь одна особая форма линейных
ускорений; естественно, макулы реагируют и на другие. Однако ускорение силы
тяжести насколько велико, что в его присутствии прочие

Рис. 12.3.
Схема левого горизонтального полукружного канала (вид сверху). За исключением
вздутия, обозначающего утрикулус, прочие части лабиринта не показаны. Угловое
ускорение в направлении, указанном черной
стрелкой (представьте себе, что вы вращаете учебник в эту сторону),
отклоняет купулу по ходу красной стрелки
линейные ускорения, встречающиеся в повседневной жизни
(например, при разгоне автомобиля), играют для вестибулярной системы
подчиненную роль и даже могут неправильно интерпретироваться ЦНС.
Естественные стимулы для полукружных
каналов. Второй вид
адекватных стимулов для ресничек вестибулярных рецепторов воспринимается в полукружных
каналах (рис. 12.3). Хотя реальная форма последних в организме–не идеальная
окружность (рис. 12.1), они действуют как замкнутые круговые трубки,
заполненные эндолимфой. В области ампулы их наружная стенка выстлана сенсорным
эпителием (рис. 12.3); здесь купула с глубоко утопленными в ней ресничками
рецепторных клеток выступает в эндолимфу. Не содержащая минеральных включений
купула полукружных каналов обладает
точно такой же плотностью, что и эндолимфа. Следовательно, линейное ускорение
(включая гравитационное) на этот орган не влияет; при прямолинейном движении и
различных ориентациях головы взаиморасположение полукружных каналов, купулы и
ресничек остается неизменным. Иной эффект у
углового (вращательного) ускорения.
При повороте головы полукружные каналы, естественно, поворачиваются вместе с
ней, однако эндолимфа в силу своей инерции в первый момент остается на месте.
Возникает разность давлений по обе стороны купулы, соединенной со стенкой
канала и образующей водонепроницаемую преграду [26], в результате чего она
отклоняется в сторону, противоположную движению (рис. 12.3). Это вызывает
сдвиговое усилие, приложенное к ресничкам, и, таким образом, изменяет
активность афферентного нерва. В горизонтальных каналах все рецепторы ориентированы
так, что киноцилии обращены к утрикулусу, поэтому активность афферентов
увеличивается, когда купула отклоняется в ту же сторону (утрикулопетально). В левом горизонтальном полукружном канале это
происходит при вращении влево. В вертикальных каналах афференты активируются
при утрикуло–фугальном отклонении
купулы (от утрикулуса). Импульсация всех этих волокон, приходящих из трех
каналов с каждой стороны, также оценивается ЦНС и дает информацию об угловых
ускорениях, действующих на голову. Именно потому, что голова может вращаться
вокруг трех пространственных осей–наклоняться вперед и назад, влево и вправо и
поворачиваться вокруг длинной оси тела, необходимы именно три полукружных
канала, лежащие в трех почти перпендикулярных друг другу плоскостях. При
вращении вокруг какой–либо диагональной оси стимулируется более одного канала.
Мозг при этом выполняет векторный анализ информации, определяя истинную ось
вращения. В клинических исследованиях
важно учитывать, что так называемый горизонтальный полукружный канал
расположен не совсем горизонтально: его передний край приподнят приблизительно
на 30°.
Особенности купулярной механики. Рассмотрим сначала, что происходит с купулой при
кратковременном угловом ускорении, т.е. когда мы просто вращаем головой. Как следует
из рис. 12.4, А, отклонение купулы соответствует не этому ускорению, а
моментальной угловой скорости. Соответственно изменения частоты нейронной
импульсации по сравнению со спонтанной приближаются к изменениям угловой
скорости, а не углового ускорения, хотя силы, вызывающие деформацию купулы,
обусловлены именно ускорением. После завершения этого короткого движения купула
возвращается в исходное состояние, и активность афферентного нерва снижается до
уровня покоя. На рис. 12.4, Б
показана принципиально иная ситуация, наблюдающаяся при длительном вращении
(например, на центрифуге), когда после первоначального ускорения надолго
устанавливается постоянная угловая скорость. Купула, отклонившись в первый
момент, затем медленно возвращается в положение покоя. Быстрая остановка
равномерного вращения снова отклоняет ее, но уже в противоположном направлении
(из–за инерции эндолимфа продолжает двигаться, в результате чего возникает
разность давлений по обе стороны купулы, приводящая к ее смещению,
характеристики которого, за исключением направленности, те же, что и в начале
движения). Для возвращения купулы в исходное положение требуется сравнительно
длительное время (10–30 с).

Рис. 12.4.
Отклонение купулы и активность афферентного нервного волокна: А–при коротком
повороте (например, головы); Б–при длительном вращении (например, на стуле).
Отметьте различие временного масштаба на рисунках
Разница между реакциями купулы на короткую и
длительную стимуляцию связана с механическими свойствами системы «купула–эндолимфа»,
которая ведет себя в первом приближении подобно сильно демпфированному
крутильному маятнику [26]. При этом следует помнить, что силы, отклоняющие
купулу, всегда обусловлены
ускорением, хотя при кратковременных угловых ускорениях, наиболее обычных в физиологических
условиях, ее отклонение пропорционально не им, а угловой скорости.
Деформации купулы, как правило, очень малы [41], но ее
рецепторы крайне чувствительны. В экспериментах на животных очень быстрый
поворот тела всего лишь на 0,005° (отклонение купулы–такого же порядка)
оказался для них надпороговым стимулом [32].
Центральная вестибулярная система
Первичные афференты вестибулярного нерва оканчиваются
главным образом в области вестибулярных ядер продолговатого мозга. С каждой
стороны тела их по четыре, отличающихся друг от друга как анатомически, так и
функционально: верхнее (Бехтерева), медиальное (Швальбе), латеральное (Дейтерса) и нижнее (Роллера). Приходящие в них
импульсы от вестибулярных рецепторов сами по себе не обеспечивают точной
информации о положении тела в пространстве, поскольку угол поворота головы
из–за подвижности шейных суставов не зависит от ориентации туловища. ЦНС должна
учитывать и положение головы относительно туловища. Следовательно,
вестибулярные ядра получают дополнительную афферентацию от шейных рецепторов (мышц и суставов) [3,21]. При экспериментальной
блокаде этих связей возникают такие же нарушения равновесия, как и при
повреждении лабиринта. В вестибулярные ядра поступают и сомато–сенсорные
сигналы от других суставов (ног, рук).
Нервные волокна, выходящие из этих ядер, связаны с
другими отделами ЦНС, что обеспечивает рефлексы поддержания равновесия. К таким
путям относятся следующие [3, 21].
а.
Вестибулоспинальный тракт, волокна
которого в конечном итоге влияют главным образом на γ–мотонейроны
мышц–разгибателей, хотя оканчиваются и на α–мотонейронах.
б. Связи с
мотонейронами шейного отдела спинного мозга, в принципе относящиеся к
вестибулоспинальному тракту.
в. Связи с
глазодвигательными ядрами, которые опосредуют движения глаз, вызываемые
вестибулярной активностью. Эти волокна проходят в составе медиального
продольного пучка.
г. Тракты, направляющиеся в вестибулярные ядра противоположной стороны мозга, дающие
возможность совместно обрабатывать афферентацию с обеих сторон тела.
д. Связи с
мозжечком, особенно с архицеребеллумом (см. ниже).
е. Связи с
ретикулярной формацией, обеспечивающие воздействие на ретикулоспинальный
тракт–еще один (полисинаптический) путь к α– и γ–мотонейронам.
ж. Тракты, проходящие через таламус в постцентральную
извилину коры головного мозга, позволяющие обрабатывать вестибулярную
информацию, а значит, ориентироваться в пространстве сознательно.
з. Волокна, направляющиеся в гипоталамус, участвующие в основном в возникновении кинетозов. Это
множество связей, лишь основные из которых перечислены выше, дают возможность
вестибулярной системе играть центральную роль в генерировании двигательной
эфферентации, обеспечивающей поддержание нужного положения тела и
соответствующие глазодвигательные реакции. При этом вертикальная поза и походка определяются главным образом
отолитовым аппаратом, тогда как полукружные каналы управляют в основном направлением взгляда. Именно
афферентация от полукружных каналов вместе с глазодвигательными механизмами
обеспечивает зрительный контакт с окружающей средой при движениях головы. При
ее вращении или наклоне глаза движутся в противоположном направлении, поэтому
изображение на сетчатке не меняется (см. статокинетические рефлексы).
Горизонтальные компенсаторные движения глаз контролируются горизонтальным
полукружным каналом, вертикальные–передним вертикальным каналом, их вращение–в
основном задним вертикальным каналом.
Еще одна важная часть ЦНС, участвующая в этих
процессах,–мозжечок, в который направляются некоторые первичные вестибулярные
афференты (так называемый прямой сенсорный мозжечковый путь) помимо вторичных,
о которых говорилось выше. Все они у млекопитающих оканчиваются в нем мшистыми
волокнами на клетках–зернах узелка (nodulus) и клочка (flocculus),
относящихся к древнему мозжечку (archicerebellum),
и частично язычка (uvula) и околоклочка (paraflocculus) старого мозжечка (paleocerebellum). Клетки–зерна оказывают возбуждающее воздействие на
клетки Пуркинье этих же областей, а аксоны последних направляются опять–таки в
вестибулярные ядра. Такая цепь осуществляет тонкую «настройку» вестибулярных
рефлексов. При дисфункции мозжечка эти рефлексы растормаживаются, что
проявляется, например, в усиленном или спонтанном нистагме (см. ниже),
нарушении равновесия, выражающемся в тенденции к падениям, неустойчивой походке
и избыточной амплитуде движений, особенно при ходьбе («петушиный шаг»).
Перечисленные симптомы относятся к синдрому мозжечковой
атаксии.
Типы импульсации нейронов вестибулярных ядер так же
разнообразны, как и их контакты, поэтому подробно мы их не рассматриваем.
Детали можно найти в специальной литературе [3, 10, 21].
Вестибулярные рефлексы; клинические тесты
Статические и статокинетические рефлексы. Равновесие поддерживается рефлекторно, без
принципиального участия в этом сознания. Выделяют статические и статокинетические рефлексы [3, 10]. Вестибулярные
рецепторы и соматосенсорные афференты, особенно от проприоцепторов шейной
области, связаны и с теми и с другими.
Статические рефлексы обеспечивают адекватное взаиморасположение
конечностей, а также устойчивую ориентацию тела в пространстве, т.е. позные рефлексы. Вестибулярная
афферентация поступает в данном случае от отолитовых органов. Статический
рефлекс, легко наблюдаемый у кошки благодаря вертикальной форме ее зрачка, –компенсаторное вращение глазного
яблока при повороте головы вокруг длинной оси тела (например, левым ухом
вниз). Зрачки при этом все время сохраняют положение, очень близкое к
вертикальному. Такой рефлекс наблюдается и у человека. Статокинетические рефлексы – это реакции на двигательные стимулы,
сами выражающиеся в движениях. Они вызываются возбуждением рецепторов
полукружных каналов и отолитовых органов; их примеры–вращение тела кошки в
падении, обеспечивающее ее приземление на все четыре лапы, или движения
человека, восстанавливающего равновесие после того, как он споткнулся.
Один из статокинетических рефлексов–вестибулярный нистагм – мы рассмотрим подробнее в
связи с его клиническим значением. Как говорилось выше, вестибулярная система
вызывает различные движения глаз; нистагм как их особая форма наблюдается в
начале более интенсивного, чем обычные короткие повороты головы, вращения. При
этом глаза поворачиваются против
направления вращения, чтобы удержать исходное изображение на сетчатке, однако,
не достигая своего крайнего возможного положения, резко «перескакивают» в
направлении вращения, и в поле зрения оказывается другой участок пространства.
Затем следует их медленное
возвратное движение.
Медленная фаза нистагма запускается вестибулярной системой,
а быстрый «перескок» взгляда–предмостовой частью ретикулярной формации.
При вращении тела вокруг вертикальной оси раздражаются
практически только горизонтальные полукружные каналы, т. е. отклонение их купул
вызывает горизонтальный нистагм. Направление
обоих его компонентов (быстрого и медленного) зависит от направления вращения
и, таким образом, от направления деформации купул. Если тело вращается вокруг
горизонтальной оси (например, проходящей через уши или саггитально через лоб),
стимулируются вертикальные полукружные каналы и возникает вертикальный, или
вращательный, нистагм. Направление нистагма принято определять по его быстрой фазе, т.е. при «правом
нистагме» взгляд «перескакивает» вправо.
При пассивном вращении тела к возникновению нистагма
ведут два фактора: стимуляция вестибулярного аппарата и перемещение поля зрения
относительно человека. Оптокинетический (вызванный зрительной афферентацией) и
вестибулярный нистагмы действуют синергически. Нейронные связи, участвующие в
этом, рассмотрены выше.
Диагностическое значение нистагма. Нистагм (обычно–так называемый «поствращательный»)
используется в клинике для тестирования
вестибулярной функции. Испытуемый сидит в специальном кресле, которое
длительное время вращается с постоянной скоростью, а затем резко
останавливается. На рис. 12.4 показано поведение при этом купулы. Остановка
вызывает ее отклонение в направлении, противоположном тому, в котором она
отклонялась в начале движения; результат–нистагм. Его направление можно
определить, регистрируя деформацию купулы; оно должно быть противоположным направлению предшествующего движения. Запись
движений глаз напоминает получаемую в случае оптокинетического нистагма (см.
рис. 11.2). Она называется
нистагмограммой.
Проведя тест на поствращательный нистагм, важно
устранить возможность фиксации взгляда в
одной точке, поскольку при глазодвигательных реакциях зрительная афферентация
доминирует над вестибулярной и в некоторых условиях способна подавить нистагм.
Поэтому испытуемому надевают очки
Френцеля с сильновыпуклыми линзами и встроенным источником света. Они
делают его «близоруким» и неспособным фиксировать взор, одновременно позволяя
врачу без труда наблюдать движения глаз. Такие очки необходимы и в тесте на
наличие спонтанного нистагма–первой,
простейшей и наиболее важной процедуре при клиническом исследовании
вестибулярной функции.
Еще один клинический способ запуска вестибулярного
нистагма – термостимуляция
горизонтальных полукружных каналов. Его преимущество–в возможности тестировать
каждую сторону тела отдельно. Голову сидящего испытуемого отклоняют назад
приблизительно на 60° (у лежащего на спине человека ее приподнимают на 30°),
чтобы горизонтальный полукружный канал занимал строго вертикальное направление.
Затем наружный слуховой проход
промывают холодной или теплой водой. Наружный край полукружного канала
расположен к нему очень близко, поэтому сразу же охлаждается или нагревается. В
соответствии с теорией Барани плотность эндолимфы при нагревании понижается;
следовательно, ее нагретая часть поднимается, создавая разность давлений по обе
стороны купулы; возникающая деформация вызывает нистагм (рис. 12.3;
изображенная ситуация соответствует нагреванию левого слухового прохода).
Исходя из его природы, этот вид нистагма называют калорическим. При нагревании он направлен к месту термического
воздействия, при охлаждении–в обратную сторону. У людей, страдающих
вестибулярными расстройствами, нистагм отличается от нормального качественно и
количественно. Детали его тестирования приведены в работе [3]. Следует
отметить, что калорический нистагм может возникать в космических кораблях в
условиях невесомости [28], когда различия плотности эндолимфы несущественны.
Следовательно, в его запуске участвует по крайней мере еще один, пока не
известный механизм, например прямое термическое воздействие на вестибулярный
орган.
Функцию отолитового аппарата можно тестировать,
наблюдая глазодвигательные реакции при наклонах головы или при
возвратно–поступательных движениях пациента, находящегося на специальной
платформе.
Нарушения вестибулярной системы. Сильные раздражения вестибулярного аппарата часто
вызывают неприятные ощущения: головокружение, рвоту, усиленное потоотделение,
тахикардию и т.д. В таких случаях говорят о
кинетозе (укачивании, «морской болезни») [23], Скорее всего это результат
воздействия комплекса необычных для организма стимулов (например, на море):
кориолисова ускорения или расхождения между зрительными и вестибулярными
сигналами. У новорожденных и больных с удаленными лабиринтами кинетозов не
наблюдается.
Для понимания причин их возникновения необходимо
учитывать, что вестибулярная система эволюционировала в условиях локомоции на
ногах, а не в расчете на ускорения, возникающие в современных самолетах.
Вследствие этого возникают сенсорные иллюзии, часто приводящие к авариям,
например, когда пилот перестает замечать вращение или его остановки,
неправильно воспринимает его направление и соответственно неадекватно
реагирует.
Острое одностороннее нарушение функции лабиринта вызывает тошноту, рвоту, потливость
и т. п., а также головокружение и иногда нистагм, направленные в здоровую
сторону. У больных наблюдается тенденция к падению в сторону с нарушенной
функцией. Очень часто, однако, клиническая картина осложнена неопределенностью
направления головокружения, нистагма и падения. При некоторых заболеваниях,
например синдроме Меньера, возникает избыточное давление эндолимфы в одном из
лабиринтов; при этом первым результатом раздражения рецепторов оказываются
симптомы, противоположные по характеру описанным выше. В противоположность
ярким проявлениям острых вестибулярных нарушений хроническое выпадение функции одного из лабиринтов компенсируется
сравнительно хорошо. Деятельность центрального отдела вестибулярной системы
может перестраиваться так, что реакция на аномальное возбуждение ослабится [21,
26], особенно когда другие сенсорные каналы, например зрительные или
тактильные, обеспечивают корректирующую афферентацию. Поэтому патологические
проявления хронических вестибулярных расстройств более выражены в темноте.
Острые двусторонние дисфункции у человека редки. В опытах на животных их
симптомы намного слабее, чем при одностороннем нарушении, поскольку
двустороннее прерывание афферентации вестибулярных ядер не затрагивает
«симметрии» организма– Невесомость (при космических полетах) не влияет на
полукружные каналы, но устраняет действие силы тяжести на отолиты, и отолитовые
мембраны во всех макулах занимают положение, определяющееся их собственными
упругими свойствами. Возникающая картина возбуждения никогда не встречается на
Земле, что может приводить к симптомам укачивания. По мере привыкания к
условиям невесомости большее значение приобретает зрительная афферентация, а
роль отолитового аппарата снижается [16].
Обиходное различие между физическим и биологическим
аспектами слуха отражается в терминологии. «Акустическими» называют физические
свойства звука, а также механические устройства или анатомические структуры, на
которые они влияют. Говоря о физиологических процессах слуха и их анатомических
коррелятах, используют термин «слуховой».
Физические свойства звукового стимула
(акустика)
Звук–это колебания молекул (Речь идет о колебаниях,
накладывающихся на броуновское движение молекул) упругой среды (в
частности, воздуха), распространяющиеся в ней в виде продольной волны давления.
Такие колебания среды генерируются колеблющимися телами, например камертоном
или раструбом громкоговорителя, которые передают ей энергию, сообщая ускорение
ближайшим к ним молекулам. От последних энергия переходит к молекулам,
расположенным чуть дальше, и т.д. Этот процесс распространяется вокруг
источника звука как волна со скоростью (в воздухе) около 335 м/с. В результате
колебания молекул в среде возникают зоны с большей или меньшей плотностью их
упаковки, где давление соответственно выше или ниже среднего. Амплитуда его
изменения называется звуковым давлением.
Его можно измерить с помощью специальных микрофонов, зарегистрировав
эффективное значение (см. учебник физики) и частотные особенности, которые и
служат характеристиками звука. Как и любое другое, звуковое давление выражают в
Н/м2 (Па), однако в акустике обычно применяют сравнительную
величину–так называемый уровень
звукового давления (УЗД), измеряемый в децибелах (дБ). Для этого
интересующее нас звуковое давление рх делят на произвольно выбранное
эталонное р0 равное 2–10–5 Н/м2 (оно близко к
пределу слышимости для человека), а десятичный логарифм частного умножают на
20. Таким образом,
УЗД =20
lg рх / рo [ДБ]
Логарифмическая шкала выбрана потому, что облегчает
описание широкого диапазона звукового давления в пределах слышимости. Множитель
20 объясняется просто: десятичный логарифм отношения силы звуков (I), исходно
названный «бел» (в честь Александра Белла), равен 10 дБ. Однако звуковое
давление р измерить легче, чем силу звука. Поскольку последняя пропорциональна
квадрату амплитуды давления (I ~ р2) и Igp2 = 2lgp, этот коэффициент введен в уравнение. Такого рода
измерения проводятся в основном в технике связи. Уровень звукового давления для
тона с давлением звука 2 • 10–1 Н/м2, например,
вычисляется следующим образом:
рх
/ рo = 2▪ 10–1/2▪
10–5=104, УЗД= 20 1g 104=20•4=80.
Таким образом, звуковое давление 2–10–1 Н/м2 соответствует
УЗД 80 дБ. Легко видеть, что удвоение звукового давления повышает УЗД на 6 дБ,
а увеличение в 10 равна 20 дБ. Ординаты на рис. 12.8 слева иллюстрируют связь
между этими параметрами.
В акустике обычно уточняют: «дБ УЗД», поскольку
дБ–шкала широко применяется для описания других явлений (например, напряжения)
или с другими условными значениями эталонов. Дополнение «УЗД» подчеркивает, что
число получено по приведенному выше уравнению с рo = 2 10–5 Н/м2.
Сила звука–это количество энергии, проходящей через единицу поверхности за
единицу времени; она выражается в Вт/м2. Величине 10–12
Вт/м2 в плоскости звуковой волны соответствует давление 2 10–5
Н/м2.
Частота звука выражается в герцах (Гц); один герц
равен одному циклу колебаний в секунду. Частота звука та же, что у его
источника, если последний неподвижен.
Звук, образованный колебаниями одной частоты,
называется тоном. На рис. 12.5, А
показана временная характеристика звукового давления для этого случая. Однако
чистые тоны в повседневной жизни практически не встречаются; большинство звуков
образовано наложением нескольких частот (рис. 12.5, Б). Обычно это сочетание основной частоты и нескольких кратных ей
по величине гармоник. Таковы музыкальные
звуки. Основная частота отражается

Рис. 12.5.
Изменение звукового давления (р) во времени: А– чистый тон; Б–
музыкальный звук; В– шум. Т– период основной музыкальной частоты;
у шума периода нет
в периоде сложной волны звукового давления (Т на рис.
12.5, Б). Так как различные источники
образуют разные гармоники, звуки при одинаковой основной частоте могут
различаться, чем и достигается богатство оттенков звучания при игре оркестра
[25]. Звук, состоящий из множества несвязанных между собой частот, называется шумом (рис. 12.5, В), в частности «белым шумом», если в нем в равной степени
представлены практически все частоты в диапазоне слышимости. Регистрируя
звуковое давление шума, периодичность обнаружить не удается.
Анатомические основы слуха;
периферический отдел органа слуха
Звуковые волны направляются в слуховую систему через наружное ухо–наружный слуховой
проход–к барабанной перепонке (рис.
12.6). Эта тонкая, с перламутровым блеском, мембрана отделяет слуховой проход
от среднего уха, в котором также
находится воздух. В полости среднего уха расположена цепочка из трех подвижно
сочлененных слуховых косточек: молоточка (malleus), наковальни
(incus) и стремени
(stapes). «Рукоятка» молоточка прочно соединена с барабанной
перепонкой, а основание стремени (которое действительно выглядит как стремя)
входит в отверстие каменистой кости овальное
окно. Здесь стремя граничит с
внутренним ухом. Энергия звука передается в него от барабанной перепонки
через колеблющиеся синхронно с ней молоточек, наковальню и стремя. Полость
среднего уха соединена с глоткой евстахиевой трубой. При

Рис. 12.6.
Схема наружного, среднего и внутреннего уха. М– молоточек, Н–наковальня,
С–стремя. Стрелки указывают соответствующие друг другу направления движения
барабанной перепонки (когда она выгнута внутрь), сочленения между наковальней и
стременем и улитковой жидкости
глотании этот проход открывается, вентилируя среднее
ухо и уравнивая давление в нем с атмосферным. При воспалительном процессе
слизистые оболочки здесь набухают, закрывая просвет трубы. Если внешнее
давление меняется (например, в самолете) или воздух из полости среднего уха
«откачивается», возникает разность давлений–«закладывает уши». Давление в этом
воздушном пространстве важно учитывать и при нырянии; ныряльщик должен
стараться, нагнетая воздух в ротовой полости («продувая уши») или делая
глотательные движения, уравнять его с возрастающим наружным давлением. Если это
не удается, возникает опасность разрыва барабанной перепонки.
Внутреннее ухо помещается в каменистой части височной
кости вместе с органом равновесия. Из–за своей формы слуховой орган назван улиткой (cochlea). Он состоит из трех параллельных свернутых вместе
каналов–барабанной (scala tympani), средней (scala media) и вестибулярной (scala
vestibuli) лестниц. Вестибулярная и барабанная лестницы соединяются между собой через геликотрему (рис. 12.6). Они заполнены перилимфой, сходной по составу с внеклеточной
жидкостью и содержащей много ионов натрия (около 140 ммоль/л [2]). Вероятно,
это–ультрафильтрат плазмы. Пространства, заполненные перилимфой и
спинномозговой жидкостью, взаимосвязаны, однако их функциональные
взаимоотношения неизвестны. Во всяком случае, спинномозговая жидкость и
перилимфа очень сходны по химическому составу.
Средняя лестница заполнена
эндолимфой. Эта жидкость богата ионами калия (приблизительно 155 ммоль/л),
т.е. напоминает внутриклеточную [2]. Пери– и эндолимфатические пространства
улитки соединены с соответствующими областями вестибулярного аппарата (рис.
12.6). Основание стремени в овальном окне примыкает к перилимфе вестибулярной
лестницы; отверстие замыкается кольцевой
связкой, так что жидкость не может просочиться в среднее ухо. Оно
сообщается с основанием барабанной лестницы еще одним отверстием–круглым окном, также закрытым тонкой
мембраной, удерживающей внутри перилимфу.
На рис. 12.7 показано поперечное сечение улитки.
Вестибулярная лестница отделена от средней
рейснеровой мембраной, а средняя от барабанной–основной (базилярной) мембраной. Утолщение, проходящее вдоль
последней,–кортиев орган – содержит рецепторы, окруженные опорными клетками.
Рецепторы–это волосковые клетки, несущие, однако, только стереоцилии; киноцилии
у них редуцированы [39]. Различают
внутренние и наружные волосковые клетки, расположенные соответственно в
один и три ряда. У человека приблизительно 3500 внутренних и 12000 наружных
волосковых клеток [12].
Как и в вестибулярном аппарате,–вторичные сенсорные клетки.
Иннервирующие их афферентные волокна отходят от биполярных клеток
расположенного в центре улитки
спирального ганглия; другие их отростки направляются в ЦНС. Около 90% нервных
волокон спирального ганглия оканчиваются на внутренних волосковых клетках,
каждая из которых образует контакты с множеством из них; остальные 10%
иннервируют значительно более многочисленные наружные волосковые клетки. Чтобы
достичь всех их, эти волокна сильно ветвятся, хотя иннервируемые одним волокном
рецепторы расположены поблизости друг от друга. Всего в слуховом нерве примерно
30000–40000 афферентных волокон [12]. К кортиеву органу подходят и эфференты,
функциональное значение которых неясно [37], хотя известно, что они могут
тормозить активность афферентов.
Над кортиевым органом лежит текториальная (покровная) мембрана–желеобразная масса, соединенная
с ним самим и с внутренней стенкой улитки. Эта мембрана отделяет узкое
заполненное жидкостью пространство под собой от эндолимфы средней лестницы
сверху. Концы стереоцилии наружных волосковых клеток связаны с нижней
поверхностью текториальной мембраны. Вероятно, реснички внутренних волосковых
клеток также с ней контактируют, хотя и значительно менее жестко; этот вопрос
окончательно еще не выяснен [39].
На наружной стороне средней лестницы расположена сосудистая полоска (stria vascularis)–область
с высокой метаболической активностью и хорошим кровоснабжением, что и
отразилось в ее названии. Она играет важную роль в обеспечении улитки энергией и регуляции состава эндолимфы.
Различные ионные насосы, включая калиевый, поддерживают постоянство ионной
среды и положительный потенциал последней [2, 20]. Известно, что некоторые
диуретические препараты (вещества, увеличивающие мочеотделение) оказывают
ототоксическое побочное действие и могут приводить к глухоте, поскольку влияют
на ионные насосы сосудистой полоски. Эти же вещества блокируют в эпителии
почечных канальцев ионные насосы (см. разд. 30.4), ответственные за реабсорбцию
солей. Очевидно, некоторые механизмы ионного транспорта в обоих случаях сходны.
Пороги слышимости. Чтобы звук был слышим, должен превышаться
определенный уровень звукового давления (УЗД). Этот порог (рис. 12.8) зависит от
частоты; человеческое ухо наиболее чувствительно в диапазоне 2000–5000 Гц. За
его пределами для достижения порога требуются значительно более высокие УЗД.

Рис. 12.7. Схема внутреннего уха в разрезе. Вверху
взаиморасположение улитки, спирального ганглия и слухового нерва.
Внизу–важнейшие элементы одного из витков спирали улитки и ее лимфатических
пространств. Состав субтекториальной лимфы точно не установлен. Тут же показаны
пространственные связи между текториальной мембраной и рецепторными клетками кортиева
органа
Громкость. Тон какой–либо частоты при превышении порога
слышимости звучит для нас громче по мере увеличения звукового давления. Связь
между физическим значением УЗД и субъективно воспринимаемой громкостью можно описать количественно. Иными словами, у человека
можно выяснить не только, слышит ли он данный тон, но и воспринимает ли он два
последовательных тона одинаковой или различной частоты как одинаково громкие или различающиеся по этому показателю. Например,
один за другим предъявляются тестируемый и эталонный тоны частотой 1 кГц, и
испытуемому предлагают отрегулировать потенциометром громкость второго звука,
чтобы она воспринималась им так
же, как предыдущая. Громкость любого звука выражают в
фонах–УЗД тона с частотой 1 кГц с равной громкостью. Таким образом, если в
приведенном выше примере субъективное ощущение уравнивается на уровне 70 дБ,
значит, громкость тестируемого тона 70 фон. Поскольку частота 1 кГц
используется как стандарт, значения в децибелах и фонах здесь одинаковые (рис. 12.8). На рис. 12.8
также приведены кривые равной слышимости, построенные по средней реакции
молодых здоровых испытуемых (крупная международная выборка). Все тоны на каждой
кривой оцениваются как одинаково громкие независимо от их частоты. Такие кривые
называют изофонами. Приведенная здесь же пороговая

Рис. 12.8.
Кривые равных уровней громкости (изофоны) в соответствии с немецким стандартом DIN 45630. На осях ординат слева отложены эквивалентные
значения звукового давления и УЗД. Красным
обозначена речевая область (см. текст)
кривая–также изофона, поскольку все ее тоны
воспринимаются как одинаково громкие, т. е. едва слышимые. Средний порог
слышимости у здорового человека равен 4 фон, хотя, конечно, возможны отклонения
от этого значения в обе стороны.
Порог различения силы звука. Так как шкала фонов основана на субъективном
восприятии, интересно установить, насколько оно точно, т.е. насколько должны
различаться звуковые давления двух тонов (которые для простоты могут иметь
одинаковую частоту), чтобы их громкость воспринималась неодинаково. В
экспериментах по измерению порога
различения силы звука эта разница оказалась очень небольшой. В области
порога слышимости два тона равной частоты воспринимаются как неодинаково
громкие, когда их УЗД различаются на 3–5 дБ. При силе звука примерно на 40 дБ
выше порога слышимости эта величина снижается до 1 дБ [49].
Шкала фонов сама по себе ничего не говорит о
субъективном увеличении громкости при
повышении УЗД. Она основана только на словах испытуемого, определяющего, когда
громкость тестируемого и эталонного тонов кажется ему одинаковой; насколько изменилась для него громкость,
в этом случае вообще не исследуется. В то же время связь между ней и звуковым
давлением представляет интерес, поскольку для оценки вредных для здоровья шумов
необходимо учитывать изменения ощущаемой громкости. Для определения этой
взаимосвязи испытуемому предлагали отрегулировать тестируемый тон частотой 1
кГц так, чтобы он казался в n раз громче
(например, в 2 или 4 раза) эталонного с той же частотой и УЗД 40 дБ. Исходя из
полученных таким образом УЗД, можно количественно описать интенсивность
ощущения; такая единица громкости называется сои. Громкость тона, звучащего для человека в 4 раза громче
стандартного, составляет 4 сон, вполовину тише–0,5 сон и т.д.
Оказалось, что при УЗД выше 30 дБ ощущение громкости
связано со звуковым давлением степенной зависимостью с показателем 0,6 при
частоте 1 кГц (степенная функция Стивенса; см. [17, 49]).
Иными словами, при частоте 1 кГц и УЗД выше 30 дБ ощущение
громкости удваивается при увеличении УЗД на 10 дБ. Поскольку удвоение звукового
давления равносильно повышению УЗД на 6 дБ, ощущение громкости параллельно не
удваивается–для этого звуковое давление должно быть почти утроено.
Следовательно, так как I ~ р2, для удвоения субъективной громкости
сила звука должна вырасти в 10 раз. Значит, громкость десяти музыкальных
инструментов, играющих в одном тоне с одинаковым УЗД, всего вдвое выше, чем у
одного из них.
Поскольку для каждого громкость в фонах по определению
выводится из звучания тона частотой 1 кГц, громкость любого тона в сонах можно
вычислить по количеству в нем фонов и кривой громкости тона 1 кГц [49]. При
технических измерениях вредных шумов применяется упрощенная процедура, дающая
приблизительные значения громкости в фонах.
Приборы для измерения УЗД и уровня
громкости. Как говорилось выше, изофоны
получены в психофизических экспериментах. Следовательно, определить громкость в
фонах физическими методами, как это делается при измерении с помощью соответствующих
микрофонов и усилителей звукового давления, невозможно. Чтобы хотя бы
приблизительно измерить уровень громкости, можно использовать такие же приборы
с частотными фильтрами, примерно соответствующими по характеристикам порогу
слышимости или другим изофонам, т. е. устройства с почти такой же неодинаковой
чувствительностью к различным частотам, как и человеческое ухо: менее
чувствительные к низким и высоким частотам. Существуют три такие международные
характеристики фильтров–А, В и С. Приводя результаты замеров, указывают, какая
из них использована, добавляя к значению в децибелах соответствующую букву,
например, 30 дБ (А), что означает приблизительно 30 фон. Характеристика фильтра
А соответствует кривой порога слышимости и должна, по идее, применяться только
при слабой силе звука, однако для простоты сейчас почти все результаты
представляются в виде дБ (А), даже если это и вносит
дополнительную ошибку. Той же шкалой пользуются при измерении вредных шумов,
хотя, строго говоря, в данном случае должна быть использована шкала сонов.
Например, шум работающей на холостом ходу автомашины составляет около 75 дБ(А).
Звуковая травма.
Если резко повысить УЗД, в конечном итоге возникнет ощущение боли в ушах. Эксперименты показали, что
для этого требуется уровень громкости около 130 фон. Более того, звук такой
силы вызывает не только боль, но и обратимую утрату слуха (временное повышение
порога слышимости) или, если воздействие было длительным,–его необратимую
утрату (стойкое повышение порога слышимости, звуковая травма). При этом
повреждаются сенсорные клетки и нарушается микроциркуляция в улитке. Звуковая
травма может возникнуть и при достаточно длительном воздействии гораздо более
слабых звуков интенсивностью не менее 90 дБ (А) [14].
Лицам, регулярно подвергающимся воздействию таких
звуков, угрожает потеря слуха; и им следует пользоваться предохранительными
приспособлениями (наушники, «беруши»). Если не принимать мер предосторожности,
в течение нескольких лет развивается тугоухость.
Субъективные реакции на шум. Помимо звуковой травмы, т. е. объективно наблюдаемого
повреждения внутреннего уха, звук может вызывать также некоторые неприятные
ощущения субъективного характера (сопровождающиеся иногда и объективными
симптомами–повышением артериального давления, бессонницей и т.д.). Дискомфорт,
вызываемый шумом, в значительной степени зависит от психологического отношения
субъекта к источнику звука. Например, жильца дома может сильно раздражать игра
на пианино двумя этажами выше, хотя уровень громкости объективно невелик и у
других жильцов жалоб не возникает. Найти общие правила, предотвращающие
неприятные для человека шумы, трудно, а действующие на этот счет
законодательные нормативы часто представляют собой лишь неудовлетворительные
компромиссы [14].
Пределы слышимости и речевая область. Слышимость тона, как показывает рис. 12.8, зависит
как от его частоты, так и от звукового давления. Молодой здоровый человек
различает частоты от 20 до 16000 Гц (16 кГц). Частоты выше 16 кГц называются ультразвуковыми, а ниже 20 Гц–инфразвуковыми. Пределы слышимости
для человека, таким образом, составляют 20 Гц–16 кГц и 4–130 фон. На рис. 12.8 зона слышимости расположена между
верхней и нижней кривыми. Частоты и силы звука, характерные для речи, находятся
в середине этой области (закрашено на рисунке красным); они соответствуют речевой зоне. Чтобы обеспечить
адекватное понимание речи, системы связи (например, телефон) должны передавать
частоты в диапазоне по крайней мере от 300 Гц до 3,5 кГц. Чувствительность к
высоким частотам с возрастом постепенно снижается (так называемая старческая тугоухость).
Порог различения частот. Из повседневного опыта известно, что тоны различаются
не только по громкости, но и по высоте,
которая коррелирует с их частотой. Тон называется высоким, если его частота
высока, и наоборот. Способность человека различать высоты последовательно
слышимых тонов поразительно высока. В оптимальной области около 1 кГц порог различения частот составляет
0,3%, т.е. около 3 Гц [14, 49].
Музыкальным звукам, включающим несколько частот, также
можно приписать определенную высоту; обычно ее считают такой же, как у чистого
тона с основной частотой звука [25]. Обычный музыкальный звукоряд делится на
октавы;
одноименные звуки соседних октав различаются по
частоте вдвое. Темперированная октава делится на 12 ступеней, каждая из которых
отличается от последующей по частоте в 1,0595 раз. Эта разница существенно
вышеупомянутого порога различения частот. Тем не менее для различения двух
одновременно звучащих чистых тонов необходима значительно большая их разница по
частоте, чем когда они следуют один за другим [17, 25, 49]. Очевидно, для этого
два стимулируемых одновременно участка внутреннего уха должны быть разделены
определенным минимальным расстоянием.
Отсюда вытекает концепция «критической частотной
полосы». Например, установлено, что слуховая система не способна различать
чистые тоны в пределах трети октавы (это и есть критическая полоса); они
сливаются, создавая ощущение одного звука. С увеличением в этом частотном
диапазоне числа составляющих звук компонентов увеличивается только субъективная
громкость, однако высота тона, воспринимаемая человеком, не меняется. Таким
образом, звуковая энергия в критической полосе суммируется, вызывая единое
ощущение.
Критическая полоса–удивительно широка: нельзя
различить два одновременно звучащих чистых
тона, разделенных почти третью октавы. В случае смешанных тонов ситуация,
конечно, иная: легко определить, когда две соседние клавиши фортепиано нажаты
одновременно, поскольку не все гармоники, накладывающиеся на основную частоту
каждой ноты, входят в единую критическую полосу.
В пределы слышимости человека укладывается около 24
критических полос. Более подробно этот вопрос рассмотрен в работах [17, 49].
Когда два тона звучат одновременно, пороги слышимости
обоих меняются. Например, на фоне постоянного тона частотой 500 Гц с УЗД 80 дБ
другие тоны с силой звука, соответствующей порогу их слышимости на рис. 12.8,
не воспринимаются. Для их слышимости необходим значительно более высокий УЗД, в
частности около 40 дБ для частоты 1 кГц [17, 49]. Это явление называется
маскированием. Оно имеет большое практическое значение, поскольку в
повседневной жизни важная акустическая информация, например разговор, может
настолько маскироваться фоновым шумом, что становится совершенно непонятной.
Более подробно психоакустические феномены описаны в работах [17, 25, 49].
Как уже говорилось, барабанная перепонка колеблется
звуком и передает его энергию в
воздушной среде по цепи косточек перилимфе вестибулярной лестницы.
Затем звук распространяется в жидкую среду внутреннего
уха; при этом большая часть его энергии отражается
от границы раздела сред, поскольку они различаются по акустическому
сопротивлению (импедансу). Однако
костнотимпанальный аппарат среднего уха «подстраивает» импедансы обеих сред
друг к другу, значительно уменьшая потери на отражение. В первом приближении
это можно сравнить с действием объектива фотокамеры, уменьшающего отражение
света на поверхности раздела «воздух–стекло». Согласование импедансов обеспечивается двумя механизмами.
Во–первых, у барабанной перепонки площадь значительно больше, чем у основания
стремени, а поскольку давление прямо пропорционально силе и обратно – площади,
оно в овальном окне выше, чем на барабанной перепонке. Во–вторых,
дополнительное увеличение давления происходит за счет изменения плеч рычага,
создаваемых цепью косточек. Таким образом, вся система действует как повышающий
электротрансформатор, хотя в процессе действуют и другие факторы–масса и
упругость связанных между собой косточек, а также кривизна и колебательные
свойства барабанной перепонки. Механизм согласования импедансов улучшает слух
на 10–20 дБ; в зависимости от частоты это эквивалентно повышению ощущаемой
громкости в 2–4 раза. Проводящие свойства тимпанально–косточкового аппарата
определяются частотой. Наилучшая передача наблюдается в середине частотного
диапазона слышимости, что отчасти обусловливает форму кривой, характеризующей
его порог.
Ощущение звука возникает и тогда, когда колеблющийся
предмет, например камертон, помещен непосредственно на череп; в этом случае
основная часть энергии передается костями последнего (так называемая костная проводимость). Как будет
показано в следующем разделе, для возбуждения рецепторов внутреннего уха необходимо
движение жидкости типа вызываемого колебаниями стремени при распространении
звука через воздушную среду. Звук, передаваемый через кости, вызывает такое
движение двумя путями. Во–первых, волны сжатия и разрежения, проходя по черепу,
вытесняют жидкость из объемистого вестибулярного лабиринта в улитку, а затем
обратно (компрессионная теория [12]). Во–вторых, масса тимпанально–косточкового
аппарата и связанная с ней инерция приводят к отставанию его колебаний от
свойственных костям черепа. В результате стремя движется относительно
каменистой кости, возбуждая внутреннее ухо (массоинерционная теория [12]).
В повседневной жизни костная проводимость не так
существенна. Разве что неузнаваемым кажется свой собственный записанный на
магнитофон голос (особенно в низкочастотном диапазоне), поскольку при живой
речи часть энергии передается в ухо по костям. Однако в диагностике костная
проводимость широко используется.
Мышцы среднего уха (m. tensor tympani, m. stapedius)
прикрепляются соответственно к молоточку и стремени. При воздействии звука их
рефлекторное сокращение ослабляет передачу, поскольку импеданс среднего уха
изменяется. Этот механизм не защищает от звуков избыточной громкости, хотя
такая возможность и обсуждалась. Функциональное значение рефлексов среднего уха
остается неясным [12].
Слуховые процессы во внутреннем ухе
Механические явления. Когда звук вызывает колебания стремени, оно передает
его энергию перилимфе вестибулярной лестницы (рис. 12.6). Поскольку жидкость во
внутреннем ухе несжимаема, должна существовать какая–то структура,
обеспечивающая
уравнивание давлений. Это–круглое окно. Его мембрана
выгибается в направлении, противоположном движению стремени. Последнее в то же
самое время выводит из состояния покоя ближайшую к нему базальную часть средней
лестницы вместе с охватывающими ее рейснеровой и основной мембранами, и она
колеблется вверх и вниз по направлению то к вестибулярной, то к барабанной
лестнице– Для простоты в дальнейшем мы будем называть среднюю лестницу вместе с
ее мебранами эндолимфатическим каналом.
Смещение его основания генерирует волну, распространяющуюся от стремени к
геликотреме, как по туго натянутой веревке. На рис. 12.9, А показаны два состояния такой волны (эндолимфатический канал
представлен одиночной линией). Поскольку звук непрерывно колеблет стремя, к
геликотреме регулярно следуют так называемые бегущие волны (см. [45]). Жесткость базилярной мембраны от
стремени к геликотреме снижается, поэтому скорость распространения волн
постепенно падает, а их длина уменьшается. По той же причине их амплитуда
сначала увеличивается (рис. 12.9), становясь значительно больше, чем около
стремени, но под действием гасящих свойств заполненных жидкостью каналов
внутреннего уха вскоре после этого уменьшается до нуля, обычно–еще перед
геликотремой. Где–то между точками возникновения волны и ее затухания находится
участок, где ее амплитуда максимальна (рис. 12.9). Этот амплитудный максимум зависит от частоты: чем она выше, тем он ближе
к стремени: чем ниже, тем дальше. В результате амплитудному максимуму каждой
частоты в диапазоне слышимости соответствует специфический участок
эндолимфатического канала (базилярной мембраны). Это называют частотной дисперсией. Сенсорные клетки
сильнее всего возбуждаются там, где амплитуда колебаний максимальна, поэтому
разные частоты действуют на различные клетки (теория места).
Описанные выше волновые движения, и в частности
положение амплитудного максимума, можно наблюдать с помощью метода Мессбауэра,
емкостного датчика или интерферометрических методов [45]. Замечательно, что
даже максимальная амплитуда волн крайне мала. Для звука на уровне порога
слышимости отклонение мембраны составляет всего лишь около 10–10 м
(приблизительно диаметр атома водорода!). Другой важный момент–строгая
локализация амплитудного максимума: различные участки базилярной мембраны очень
четко «настроены» на определенную частоту, если улитка полностью интактна. При
ее повреждении (например, при легкой гипоксии) амплитуда колебаний снижается, и
такая тонкая настройка утрачивается. Другими словами, базилярная мембрана не
просто пассивно колеблется; активные процессы обеспечивает частотноспецифичный
механизм усиления.
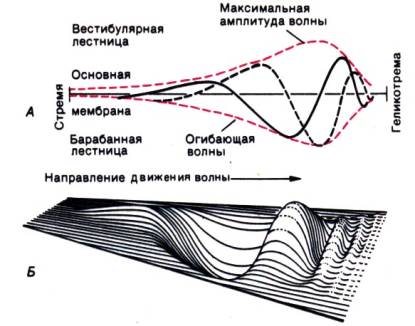
Рис. 12.9. А. Схема бегущей волны в два момента времени.
Огибающая показывает ее максимальную амплитуду при постоянной частоте в
различных участках улитки. Б.
Трехмерная реконструкция волны
Процессы преобразования в волосковых
клетках. Как
говорилось в предыдущем разделе, благодаря механическим свойствам улитки
определенная звуковая частота вызывает колебания базилярной мембраны с
достаточной для возбуждения сенсорных клеток амплитудой только в одном, строго
ограниченном месте. Поскольку базилярная и текториальная мембраны движутся относительно друг друга, возникает
действующее на реснички сдвиговое усилие–как при прямом их контакте с
текториальной мембраной, так и в результате перемещения субтекториальной лимфы;
в обоих случаях их изгибание служит адекватным стимулом для слуховых рецепторов
(как у вестибулярных рецепторов).
Это изгибание запускает процесс преобразования (трансдукции): микроскопические механические
деформации ресничек приводят к открытию ионных каналов в мембране волосковых
клеток и, следовательно, к их деполяризации. Ее предпосылкой служит наличие эндокохлеарного потенциала.
Микроэлектродные измерения показали, что у эндолимфатического пространства
положительный (приблизительно + 80 мВ) заряд относительно вестибулярной
лестницы и других внеклеточных пространств организма. Сосудистая полоска и
кортиев орган несут отрицательный заряд (~ –70 мВ; рис. 12.10). Потенциалы,
регистрируемые в кортиевом органе, вероятно, соответствуют внутриклеточным
потенциалам волосковых и опорных клеток. Положительный эндокохлеарный потенциал
обеспечивается энергозависимыми процессами в сосудистой .полоске. Сдвиг
ресничек при стимуляции изменяет сопротивление мембраны волосковых клеток в
результате открытия ионных каналов. Поскольку между

Рис. 12.10.
Постоянные потенциалы улитки

Рис. 12.11.
Микрофонный потенциал улитки (МП) и составной потенциал действия (СПД)
слухового нерва, зарегистрированные у круглого окна при звуке щелчка
эндолимфатическим пространством и их внутриклеточной
средой существует значительная разность потенциалов (не менее 150 мВ),
синхронно со стимулом возникают локальные ионные токи, меняющие мембранный
потенциал волосковых клеток, т. е. генерирующие рецепторный потенциал (так
называемая гипотеза батареи) [7, 29,
48]. Зарегистрировать его трудно, но возможно [7, 29, 48]. Проще все же, поместив
микроэлектроды вблизи рецепторов в барабанную лестницу или на круглое окно,
записать микрофонный потенциал улитки
(рис. 12.11).
Он аналогичен выходному напряжению микрофона и
достаточно точно отражает изменения звукового давления. Магнитофонная запись речи,
выполненная путем подключения к микрофонному потенциалу подопытного животного,
вполне разборчива. Происхождение этого потенциала неясно; первоначальное
предположение о том, что он состоит из регистрируемых внеклеточно компонентов
рецепторных потенциалов волосковых клеток, уже не вполне приемлемо. Как
показали внутриклеточные отведения из внутренних и наружных волосковых клеток
[7, 33, 34], хотя рецепторные потенциалы ими и генерируются, при высокой
частоте стимулов регистрируется только постоянное напряжение: мембранный
потенциал волосковых клеток не меняется синхронно с высокочастотным звуком.
Микрофонный же потенциал:
1) синхронен звуковому стимулу практически без
латентного периода;
2) лишен рефракторного периода;
3) лишен измеримого порога;
4) не подвержен утомлению; т.е. во всех отношениях
отличается от нейронного потенциала действия.
Деполяризация волосковых клеток вызывает выброс из их
базальной части медиатора (возможно, глутамата [38]), возбуждающего афферентные
нервные волокна. Когда около уха раздается щелчок (короткий импульс давления),
волокна слухового нерва активируются синхронно и от круглого окна, помимо
микрофонного, можно записать еще и составной потенциал действия. Более
длительные звуки вызывают асинхронную импульсацию, не суммирующуюся в отдельные
потенциалы действия. На рис. 12.11 показаны микрофонный потенциал улитки (МПУ)
и составной потенциал действия (СПД), вызванные щелчком. Они были
зарегистрированы на кошках, однако их можно записать и у человека, когда в
диагностических целях электрод пропускают через барабанную перепонку и подводят
к круглому окну.
Кодирование звука в волокнах слухового
нерва.
В улитковом нерве 90% афферентных волокон
миелинизированы и отходят от внутренних волосковых клеток. Каждое контактирует
только с одной из них, т.е. с очень малой частью улитки. Эти волокна достаточно
толсты для регистрации в них с помощью микроэлектродов потенциалов действия и
изучения ответа на звуковую стимуляцию (волокна, отходящие от наружных
волосковых клеток, для этого слишком тонки). Так как каждый участок улитки
соответствует определенной частоте, каждое из таких волокон сильнее всего
возбуждается своей характеристической
частотой звука, а другими частотами не активируется совсем или активируется
лишь при повышении звукового давления. Это отражено на рис. 12.12, где
представлен график зависимости порога восприятия от частоты стимула для двух
разных волокон. Критерием для установления порога служит определенное повышение
активности над ее спонтанным уровнем. Каждое волокно возбуждается при значениях
частоты и интенсивности в пределах закрашенной на рисунке области. У
ограничивающей ее кривой
частотно–пороговой характеристики заметен узкий, заостренный низкопороговый
участок и широкий высокопороговый. Частотно–пороговые характеристики отражают

Рис. 12.12.
Схематичные частотно–пороговые кривые двух афферентных волокон слухового нерва
(а, б) с различными характеристическими частотами (ХЧ). Кривая в типична для
волокна с патологическими изменениями, вызванными повреждением внутреннего уха
распределение частотных максимумов на основной
мембране. Реакция одиночного волокна на стимул, выраженная в виде такой кривой,
представляет собой спектральный анализ звука. Если в нем несколько различных
частот, активируется несколько групп нервных волокон. Длительность звукового стимула кодируется продолжительностью
нервной активности, а интенсивность–ее
уровнем. При увеличении звукового давления возрастает и частота импульсации
нейронов (до некоторого предела, после чего наступает насыщение). При очень
высоком давлении, кроме того, активируются и соседние волокна, находившиеся
прежде в состоянии покоя. Такой процесс показан на рис. 12.12; оба волокна
возбуждаются, если звук соответствует области перекрывания их
частотно–пороговых характеристик. Итак, на уровне первичных афферентов звуковой
стимул раскладывается на частотные компоненты. Каждый из них возбуждает
соответствующие нервные волокна. На более высоких уровнях слухового тракта
нейроны могут вести себя и по–иному.
При повреждениях улитки чувствительность и частотная
избирательность афферентных волокон снижаются (рис. 12.12). Рецепторный
потенциал внутренних волосковых клеток также меняется и, как говорилось выше,
то же происходит с механическими колебательными свойствами основной мембраны.
Последние, можно полагать, и определяют поведение этих клеток и волокон, однако
сами зависят от процесса активного механического усиления, за который,
возможно, отвечают наружные волосковые клетки. Согласно существующей гипотезе,
они стимулируются звуком первыми и генерируют дополнительную колебательную
энергию той же частоты. Затем она передается внутренним волосковым клеткам.
Если это верно, речь идет о своего рода гибриде между сенсорной клеткой и
генератором механической энергии [7, 44].
Множество вопросов здесь пока не находит ответа,
однако способность улитки как вырабатывать, так и анализировать звуковую
энергию бесспорна. Звук, генерируемый в улитке, можно измерить даже снаружи от
барабанной перепонки [7, 36]. Эти процессы очень часто нарушаются при различных
повреждениях.
Наружные волосковые клетки подходят на роль активных
усилителей, поскольку содержат сократительные белки и обслуживаются чрезвычайно
развитой эфферентной сетью. Кроме того, их афференты явно не существенны для
передачи информации в мозг [7, 13].
Кодирование звуковых частот по принципу локализации
рецептора рассмотрено выше. Второй вид кодирования информации в слуховом нерве
следующий. Тоны частотой до 5 кГц обычно вызывают нейронную импульсацию в слуховом
нерве только в определенные фазы цикла звукового колебания. В результате
временная структура стимула (например, период Т на рис. 12.5, Б) представлена группами потенциалов
действия, передаваемых в ЦНС по слуховому нерву в соответствующие моменты времени.
Мозг, очевидно, способен оценивать временную структуру импульсации и определять
лежащую в его основе звуковую частоту (так называемый анализ периодичности). Особенно четкие доказательства этого были
получены при прямой электростимуляции слухового нерва пациентов, страдающих
глухотой; периодические стимулы перерабатывались таким образом, что возникало
ощущение тона определенной высоты, что показывает действительную важность
анализа периодичности для слуха [30, 46].
Анатомия слухового тракта
в виде сильно упрощенной схемы
показана на рис. 12.13. Для простоты изображен только путь от левого уха.
Стрелки означают синапсы с нейронами более высоких порядков. Чтобы не
перегружать рисунок, возвратные коллатерали и вставочные нейроны опущены, хотя
в слуховой системе такого рода связи весьма распространены.
Первичное афферентное волокно раздваивается, посылая
один отросток к вентральному, а
другой–к дорсальному улитковым
(кохлеарным) ядрам. Их тонкая структура (особенно–дорсальных) очень сложна
[11, 12, 40]. Вентральный тракт (из вентрального ядра) направляется (частично
через ядро трапециевидного тела) к ипси– и контралатеральному оливарным
комплексам, нейроны которых, таким образом, получают сигналы от обоих ушей.
Именно данный нейронный уровень позволяет сравнивать акустические сигналы,
поступающие с двух сторон организма (ниже мы вернемся к этому процессу
сравнения). Дорсальный тракт (из

Рис. 12.13.
Сильно упрощенная схема слухового тракта (только для левого уха). Чтобы
продемонстрировать бинауральные взаимодействия в верхней оливе, показаны также
связи правого вентрального улиткового
ядра. Центробежные пути опущены
дорсального ядра) переходит на противоположную сторону
тела и направляется в ядро латерального
лемниска (латеральной петли). Восходящие отростки клеток оливарного
комплекса как ипси–, так и контралатеральные. После синаптического переключения
в ядре латеральной петли слуховой тракт проходит через нижние холмики четверохолмия и
медиальное коленчатое тело в первичную слуховую кору, покрывающую
поперечные височные извилины верхней части височных долей (извилина Гешля). Эта
зона соответствует полю 41 по Бродману; большая его часть скрыта в глубине
сильвиевой борозды. С первичной слуховой корой соседствуют другие проекционные
участки слуховой системы, называемые вторичной слуховой корой (поле 42 по
Бродману). Таким образом, предкорковый слуховой тракт состоит по крайней мере
из пяти или шести нейронов, а поскольку дополнительные синаптические
переключения и возвратные коллатерали на рис. 12.13 не показаны, возможны и
более длинные цепи. Подробнее это изложено в работах [12, 20]. Наконец, кроме
афферентных путей в состав слуховой системы входят и центробежные эфферентные
волокна, также не представленные на рис. 12.13 [37].
Возбуждение центральных нейронов
слуховой системы. В то время
как первичные афференты слухового нерва возбуждаются чистыми тонами, т. е.
очень простыми звуковыми стимулами, нейроны более высоких уровней в основном к
этому не способны. В вентральном
улитковом ядре они еще ведут себя подобно первичным нейронам. Чистые

Рис. 12.14.
Активность четырех нейронов дорсального улиткового ядра в ответ на действие
тона характеристической частоты в течение 50 мс (по [27] с изменениями). По оси абсцисс–время; по оси ординат– число потенциалов действия
тоны надпороговой интенсивности всегда вызывают их
возбуждение; у них узкие заостренные частотно–пороговые кривые и короткие
латентные периоды. Однако уже в
дорсальном улитковом ядре картина совершенно иная [11, 12, 47]. Хотя и
здесь большинство нейронов возбуждаются чистыми тонами, типы их ответов широко
варьируют. В качестве примера на рис. 12.14 показаны реакции различных волокон,
исходящих из этого ядра: в каждом случае предъявлялся тон длительностью 50 мс с
характеристической для данной клетки частотой. Нейрон на рис. 12.14, А ведет себя подобно первичному
афференту, тогда как поведение остальных существенно иное. В некоторых из них
звук может вызывать торможение; другие возбуждаются строго определенными
частотами и тормозятся при незначительном отклонении от них. Есть и нейроны,
особым образом реагирующие на звуки переменной частоты (так называемые
частотно–модулированные тоны), хотя отвечающие и на чистые тоны. Анатомическая
основа такого сложного поведения –коллатеральные связи, одни из которых
возбуждающие, а другие тормозные.
Функциональное значение всего этого, очевидно, в том, что нейроны особенно
четко реагируют на определенные особенности звукового стимула, способствуя распознаванию
образа уже на таком низшем уровне тракта. На более высоких уровнях
специфичность их ответа постепенно возрастает.
Чем дальше от улитки по слуховому тракту, тем более
сложные звуковые характеристики требуются для активации нейронов [20]. Многие клетки
вообще не отвечают на чистые тоны. В нижних
холмиках четверохолмия, например, есть клетки, реагирующие только на
частотно–модулированные тоны со специфическими направлением и степенью
модуляции. Другие нейроны здесь отвечают только на амплитудно–модулированные
(т.е. с переменной интенсивностью) тоны. И в этом случае модуляция часто должна
обладать определенными особенностями, иначе возбуждения не вызовет.
В общем можно сказать, что информация, содержащаяся в
звуковом стимуле, многократно перекодируется по мере прохождения через
различные уровни слухового тракта. В ходе этого процесса нейроны того или иного
типа выделяют «свои» свойства стимула, что обеспечивает довольно специфичную
активацию нейронов высших уровней.
В повседневной жизни мы практически не сталкиваемся с
чистыми тонами. Окружающие нас звуки состоят из различных частотных
компонентов, которые постоянно и независимо друг от друга изменяются. Варьирует
также их амплитуда и длительность; они могут возникать и заканчиваться внезапно
или постепенно, повторяться или быть уникальными; их источник может
располагаться ближе или дальше от нас, двигаться и т. д. Человек, по крайней
мере с тренированным слухом, способен оценить все эти свойства. Нейронные
процессы, лежащие в основе такой оценки, выявлены главным образом в слуховой коре [47]. Например,
некоторые нейроны первичной слуховой коры отвечают только на начало звукового
стимула, другие–только на его окончание. Одни группы нейронов возбуждаются при
звуках определенной длительности, другие–при повторяющихся звуках. Есть и
клетки, активирующиеся только при той или иной частотной или амплитудной
модуляции звука. Многие нейроны активируются широким диапазоном частот, т.е.
шумами, у других частотно–пороговые характеристики отличаются одним или несколькими
резко выраженными минимумами. Большинство корковых клеток возбуждается
афферентами контралатерального уха, однако некоторые отвечают на
ипсилатеральную стимуляцию, а остальные–только на двустороннюю. Значительная
часть нейронов первичной слуховой коры не активируется ни при каких
экспериментальных воздействиях; возможно, они высокоспецифичны и реагируют
только на стимулы, которые слишком сложно воспроизвести в лабораторных условиях
[47].
В целом ответы клеток первичной слуховой коры сходны с известными для сложных или
сверхсложных нейронов зрительной коры. Очевидно, они участвуют в распознавании
слуховых образов–процессе, очень существенном, например, для понимания речи. Даже в слуховой коре обезьян обнаружены клетки,
отвечающие главным образом на звуки, связанные с внутривидовой коммуникацией.
Однако свойства этих нейронов часто зависят от каких–то неизвестных параметров,
и их реакции непредсказуемо варьируют [42].
Повреждение височных долей мозга, где располагается
слуховая кора, затрудняет понимание речи, пространственную локализацию
источника звука (см. ниже) и идентификацию его временных характеристик. Однако
подобные поражения не влияют на способность различать частоту и силу звука.
Более подробно центральная обработка звуковой информации рассмотрена в работах
[11, 12, 20, 47].
Недавние исследования показали, что тонотопическая
организация, свойственная улитке, сохраняется и на более высоких уровнях
слуховой системы, включая кору. Наличие такой организации, т.е. упорядоченного
распределения участков, связанных с определенными звуковыми частотами, в
первичной слуховой коре раньше отрицалось.
Другим результатом, противоречащим более ранним
предположениям, стал тот факт, что для слуховых нейронов высших уровней не
характерны выраженные пики частотно–пороговых характеристик. У первичных
афферентов слухового нерва, если экспериментальное животное находится в
оптимальных условиях, они очень четкие.
Слуховая ориентация в пространстве. Центральная слуховая система очень важна для пространственной ориентации. Как
известно из повседневного опыта, при
бинауральном слухе направление на источник звука можно определить
достаточно точно. Физическая основа такой
дирекциональности в том, что обычно одно ухо расположено от него дальше,
чем другое. Распространяясь с конечной скоростью, звук достигает более
удаленного уха позже и с меньшей силой, а слуховая система способна
выявить ее разницу в двух ушах уже на уровне 1 дБ [5, 11, 13, 43]. На рис.
12.15 показан метод расчета разницы во времени прохождения звука. Разница в расстоянии
ΔS=d sin α, где (d–расстояние между ушами, а α –угол, под которым источник звука расположен
относительно испытуемого. Значит, временная задержка Δt= Δs/с, где
с–скорость звука. Человек способен уловить задержку всего в 3–10–5
с, что соответствует отклонению источника звука от средней линии примерно на
3°. При оптимальных условиях можно различить и вдвое меньший угол.
И психофизические, и нейрофизиологические эксперименты
показали, что дирекциональный слух основан на разнице во времени проведения и силе звука. При использовании
наушников для независимой стимуляции каждого уха задержка сигнала или снижение
его интенсивности с одной из сторон вызывает ощущение локализации звука в
противоположном ухе. Задержку можно компенсировать повышением интенсивности; в
этом случае кажется, что источник звука расположен в голове [5]. Сходные
результаты получены в нейрофизиологических экспериментах. В верхней оливе,
первом уровне

Рис. 12.15.
Расчет разницы во времени достижения звуком правого и левого уха (см. текст)
слуховой системы с двусторонней афферентацией, есть
нейроны, которые по отношению к временным характеристикам и интенсивности
сигнала ведут себя сходным образом [11, 13, 42, 43]. Возбуждение в них
максимально, когда звук в одном ухе громче, чем в другом, и предшествует ему.
Другой тип клеток здесь наиболее активен, когда стимулы, достигающие обоих
ушей, определенным образом различаются по времени поступления и интенсивности.
Значит, первый тип клеток максимально реагирует на звук, локализованный по оси одного
из ушей, а другой–на приходящий под определенным углом [11, 13, 42]. В верхних
холмиках четверохолмия слуховая и зрительная афферентации, сочетаясь, дают
трехмерную «карту» пространства. В
слуховой коре некоторые клетки также активируются только при вполне
определенном расположении относительно слушателя источника звука. При ее
разрушении страдает и пространственная ориентация. Однако до сих пор не вполне
ясно, каким образом ЦНС справляется с определением временной разницы менее 10 –4
с.
Различий во времени проведения и интенсивности
недостаточно для того, чтобы понять, спереди или сзади, сверху или снизу
относительно головы находится источник звука. Для этого необходимо
дополнительное устройство–ушная раковина. Ее строение так «искажает» сигнал в
зависимости от расположения его источника, что его удается локализовать. В
технике это можно использовать, поместив в голову манекена микрофоны на место
барабанных перепонок: у полученных с их помощью стереофонических записей будет
превосходное качество [5].
Слух в условиях шума. Бинауральный слух имеет и иную, более важную, чем
ориентация в пространстве, функцию; он помогает анализировать акустическую
информацию в присутствии посторонних шумов. «Межушные» различия в интенсивности
и направлении поступления сигналов используются ЦНС для подавления фонового
шума и выделения полезных звуков (например, когда сосредоточиваешься на нужном
разговоре в многолюдном собрании). Такой селективный фильтрующий процесс
усиливает слышимость приблизительно на 10 дБ [13]. У глухих на одно ухо этого
не происходит, в чем легко убедиться, заткнув себе ухо. Следовательно, при
тугоухости важно восстанавливать бинауральный слух, например, с помощью
слуховых аппаратов.
Адаптация слуховой системы. Слуховая система, как и другие сенсорные системы,
способна к адаптации. В этом
процессе участвуют как периферическое ухо, так и центральные нейроны. Адаптация
проявляется во временном повышении слухового порога. Это полезно, поскольку
уменьшает порог различения громкости и, таким образом, способствует
дифференцировке слуховых ощущений. У адаптированного уха изофоны смещены вверх
и сближены. Более подробная информация содержится в работах [14, 49].
Патофизиология нарушения слуха
Тугоухость и глухота весьма существенно сказываются на
жизни больных, и потому привлекают большое внимание клиницистов. Причины этих
нарушений можно разделить на три категории.
1. Нарушения проведения звука. К ним относятся повреждения среднего уха. Например,
при его воспалении тимпанально–косточковый аппарат не передает нормального
количества звуковой энергии внутреннему уху. В результате, даже если оно
здорово, слух ухудшается. Существуют микрохирургические способы эффективного
устранения таких дефектов слуха.
2. Нарушения восприятия звука. В этом случае повреждены волосковые клетки кортиева
органа, так что нарушено либо преобразование сигнала, либо выделение
нейромедиатора. В результате страдает передача информации из улитки в ЦНС.
3. Ретрокохлеарные нарушения. Внутреннее и среднее ухо здоровы, но поражены либо
центральная часть первичных афферентных волокон, либо другие компоненты
слухового тракта (например, при опухоли мозга).
Проверка слуха у пациентов называется аудиометрией. Для выявления и
локализации повреждений слухового аппарата разработаны многочисленные тесты
(подробнее–см. [24]).
Важнейший среди
них–пороговая аудиометрия. Больному через один наушник предъявляются
различные тоны. Врач начинает с явно подпороговой силы звука и постепенно
увеличивает звуковое давление, пока. больной не сообщит, что слышит звук. Это
звуковое давление наносится на график (рис. 12.16), называемый аудиограммой.
На стандартных аудиографических бланках уровень
нормального порога слышимости представлен жирной чертой и помечен «О дБ». В
отличие от рис. 12.18, более высокие пороги наносятся ниже нулевой линии и
характеризуют степень утраты слуха–на сколько децибелов они ниже нормы.
Подчеркнем, что речь здесь идет не о децибелах УЗД, а об утрате слуха на
столько–то дБ. Например, если заткнуть пальцами оба уха, его снижение составит
приблизительно 20 дБ (при выполнении этого эксперимента, конечно, надо
постараться не производить слишком много шума самими пальцами). С помощью
наушников тестируется восприятие звука при его воздушной проводимости. Костную проводимость проверяют сходным
образом, но вместо наушников используется камертон, приложенный с тестируемой
стороны к сосцевидному отростку височной кости, чтобы колебания
распространялись непосредственно через череп. Сравнивая пороговые кривые для
костной и воздушной проводимости, можно различить глухоту, вызванную
повреждением среднего или внутреннего уха.
Глухота при повреждении среднего уха
обусловлена нарушением проведения звука.
Внутреннее ухо при этом здорово. В таких условиях снижение слуха выявляется в
тесте на воздушную проводимость (ср. рис. 12.16), а порог для костной
проводимости нормален, поскольку звуковая энергия при его проверке достигает
волосковых клеток, минуя среднее ухо.
Глухота при патологии внутреннего уха обусловлена повреждением волосковых клеток; среднее
ухо здорово. В этом случае порог для обоих видов проводимости повышен,
поскольку и в том, и в другом случае преобразование сигнала осуществляется
посредством одного и того же рецепторного процесса. Ретрокохлеарные нарушения
также повышают оба порога.
С помощью камертона (обычно с частотой 256 Гц)
нарушения проведения очень легко отличить от повреждений внутреннего уха или
ретрокохлеарных патологий, когда известно, какое ухо слышит хуже (тест Вебера). Ножку колеблющегося
камертона помещают на среднюю линию черепа; если поражено внутреннее ухо,
больной сообщает, что тон звучит со здоровой стороны; если среднее–с
пораженной.
Этот феномен просто объяснить в случае патологии
внутреннего уха. Поврежденные рецепторы слабее возбуждают слуховой нерв,
поэтому тон звучит громче в здоровом ухе, а за счет этого различия и возникает
дирекциональное ощущение. В случае поражения среднего уха мы сталкиваемся с
тремя одновременными процессами. Во–первых, ухудшение колебательных свойств
косточкового аппарата ослабляет передачу звука не только снаружи внутрь, но и в
противоположном направлении. Следовательно, внутреннее ухо, возбуждаясь звуком,
при





