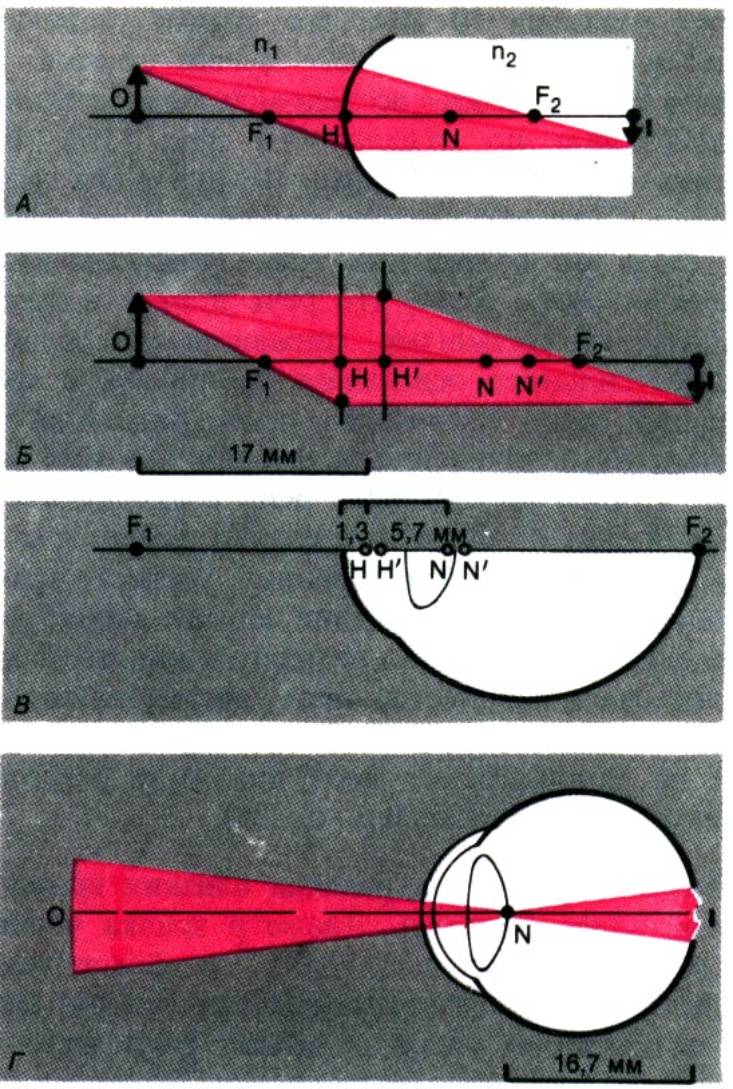
Редуцированный глаз.
Еще проще схема редуцированного глаза (рис. 11.6,В), в которой Н совпадает с
Н', a N–c N'. Здесь
расстояние от узловой точки N до сетчатки составляет 16,67 мм. Зная эту
величину и угол а, под которым виден объект, можно определить размер его
изображения на сетчатке.
Процессы регуляции в диоптрическом
аппарате
Преломляющая способность хрусталика и диаметр зрачка
изменяются при сокращении гладких мышц глаза, которые управляются нейронными
механизмами.
Реакция зрачка. В норме зрачки обоих глаз круглые и их диаметры
одинаковы. Средний диаметр зрачка уменьшается с возрастом.
Реакция на
свет. При
постоянном освещении количество света, попадающее в глаз за единицу времени,
пропорционально площади зрачка. При
снижении внешней освещенности зрачок рефлекторно расширяется. Если при дневном
свете человек закроет глаза на 10–20 с, его зрачки увеличатся. Когда он снова
их откроет, зрачки сузятся. Эту реакцию
на свет можно исследовать более детально, освещая глаза по отдельности
(рис. 11.7). Если осветить один глаз, то через 0,3–0,8 с его зрачок сократится (прямая реакция на свет); у
неосвещенного глаза он сократится тоже
(содружественная реакция на свет). Ясно, что речь идет о полезном
регуляторном механизме, снижающем в условиях слишком сильного освещения
(например, в яркий солнечный день) количество света, падающего на сетчатку, и
увеличивающем его при плохом освещении. В этой регуляторной цепи с отрицательной обратной связью датчиками служат
рецепторы сетчатки, а регулируемой переменной–диаметр зрачка У молодых людей
последний может варьировав в пределах от 1,5 мм до примерно 8 мм, что позволяет
изменять количество света, достигающего сетчатки, приблизительно в 30 раз.
Однако этот механизм не компенсирует весь диапазон колебаний внешней
освещенности.
Реакция при рассматривании близких
предметов (конвергентная реакция). Диаметр зрачка человека зависит также от расстояния до фиксируемого предмета.
Если испытуемый сначала смотрит вдаль, а затем переводит взгляд на объект,
расположенный в 30 см от него, зрачки сужаются. Поскольку оси глаз при этом
обычно сводятся друг к другу, такая реакция называется конвергентной. Настройка зрачка на ближний объект сопровождается
увеличением преломляющей силы хрусталика (см. ниже). Как и в фотоаппарате, при
уменьшении апертуры глубина резкости в глазу увеличивается.
Функция зрачковых мышц и их иннервация (рис. 11.8). Зрачковые
реакции осуществляются с помощью двух систем гладких мышц в радужной оболочке.
При сокращении кольцевой мышцы–сфинктера
зрачок сужается (миоз); при сокращении мышцы–дилататора,
волокна которой проходят в радужной оболочке радиально, он расширяется (мидриаз). Сфинктер иннервируется
парасимпатическими нервными волокнами, выходящими из цилиарного (ресничного)
ганглия, расположенного позади глаза. Преганглионарные волокна отходят от зрачководвигательных нейронов ядра
Эдингера–Вестфаля, которое является «вегетативной» частью глазодвигательного
ядра ствола мозга, и направляются к глазнице в составе глазодвигательного
нерва. Уровень активации зрачководвигательных нейронов этого ядра регулируется
нейронами претектальной зоны (рис. 11.8). Здесь оканчиваются аксоны слоя
ганглиозных клеток сетчатки и зрительной коры (полей 18 и 19). Дилататор,
напротив, иннервируется симпатическими нервными волокнами, возбуждаемыми
нейронами цилиоспинального центра,
расположенного на уровне восьмого шейного и первого, второго грудных сегментов
спинного мозга. Аксоны нейронов этого центра идут через шейный отдел
симпатической цепочки в верхний шейный ганглий, где образуют синапсы с
постганглионарными нейронами. Аксоны последних направляются в глазницу вдоль внутренней сонной и глазной артерий и переходят там в цилиарный нерв. Уровень активации

Рис. 11.7.
Схема зрачковых рефлексов: прямая и содружественная реакции на свет и конвергентная
реакция. Стрелками показано освещение одного глаза.

Рис. 11.8.
Схема иннервации мышц радужной оболочки и цилиарной мышцы. Красным показаны эфференты симпатической нервной системы, черным парасимпатической
цилиоспинального центра зависит от общего тонуса
вегетативной нервной системы.
Клиническое значение зрачковых реакций. Диаметр зрачка и зрачковые реакции важные
диагностические признаки, поскольку по ним можно выявить поражения сетчатки,
зрительного нерва, ствола мозга (глазодвигательной зоны), шейного отдела
спинного мозга, а также областей, через которые проходят пре– и
постганглионарные зрачководвигательные волокна (глубинных слоев шеи,
клиновидной кости и глазницы). Вегетативной
иннервацией мышц радужной оболочки объясняется также зависимость размера зрачка
от возраста, психологических факторов, уровня внимания и степени утомления [12,
30].
Аккомодация. Настройка преломляющей силы диоптрического аппарата
глаза человека на определенное расстояние до фиксируемого объекта (аккомодация) осуществляется за счет
изменения кривизны хрусталика, особенно его передней
поверхности. Эта кривизна зависит от его упругости и сил, воздействующих на
его сумку. Пассивные упругие силы, развиваемые цилиарным аппаратом, сосудистой
оболочкой и склерой, передаются сумке хрусталика через волокна цилиарного (ресничного, или циннова) пояска. Механическое
напряжение склеры зависит в свою очередь главным образом от внутриглазного
давления. Когда натяжение волокон пояска увеличивается, хрусталик растягивается
и, следовательно, уплощается. Влияние на него этих пассивных упругих сил может
изменяться под действием окружающей его цилиарной (ресничной) мышцы (рис. 11.5,
11.8). Волокна этой гладкой мышцы ориентированы по–разному радиально, по
окружности и сверху вниз. Они иннервируются вегетативными, в основном
парасимпатическими нервными волокнами. Когда цилиарная мышца сокращается, она
противодействует упругим силам, действующим на хрусталик через циннов поясок,
при этом натяжение сумки хрусталика уменьшается, кривизна его передней
поверхности увеличивается и преломляющая сила возрастает: происходит аккомодация. Когда цилиарная мышца
расслабляется, кривизна хрусталика и его преломляющая сила уменьшаются. В
максимально неаккомодированном состоянии здоровый глаз дает на сетчатке четкое
изображение лишь тех предметов, которые удалены на очень большое расстояние (в
«бесконечность»).
Диапазоном аккомодации называется интервал изменения преломляющей силы (в
диоптриях) при фиксации объекта, приближающегося из бесконечности на максимально
близкое расстояние четкого видения. Самый широкий диапазон в молодом возрасте
(14 диоптрий). Из уравнений (3) и (5) следует, что максимально аккомодированный
глаз способен обеспечить четкое изображение на сетчатке предметов, удаленных от
него на 1/14 м, т.е. на 7 см. С возрастом за счет некоторого обезвоживания
хрусталик постепенно снижает свою упругость, а значит, и способность изменять
преломляющую силу (диапазон аккомодации). Ближайшая точка четкого видения
постепенно отодвигается от глаза, поэтому пожилым людям для чтения обычно
требуются очки (пресбиопия).
Нервная регуляция аккомодации. Как и в зрачководвигательной системе, за аккомодацию
отвечают преганглионарные парасимпатические аксоны (см. разд. 16.1) нейронов
ядра Эдингера Вестфаля, проходящие через цилиарный ганглий. Адекватный стимул
для изменения аккомодации–нечеткость изображения
на сетчатке. Ее, по–видимому, регистрируют нейроны фовеальной проекционной
зоны зрительной коры (поле 18), связанной с ядром Эдингера–Вестфаля (рис.
11.8).
Некоторыми
лекарственными препаратами
можно воздействовать на периферические вегетативные синапсы цилиарной мышцы и
мышц радужной оболочки (как и на любые другие в вегетативной нервной системе).
Если, например, в конъюнктивальный мешок (за веко) закапать раствор атропина, он диффундирует в радужную
оболочку и цилиарное тело и блокирует передачу сигналов через парасимпатические
синапсы. В результате хрусталик станет неаккомодированным, а зрачок расширится.
И наоборот, ингибитор ацетилхолинэстеразы
неостигмин вызывает сужение зрачка и усиливает аккомодацию [12, 20, 26,
30].
Оптические недостатки глаза и аномалии
рефракции
Система линз современного фотоаппарата дает
изображение значительно более высокого качества, чем диоптрический аппарат
глаза. Герман фон Гельмгольц (1821–1894), бывший одновременно физиологом и
физиком, однажды пошутил, что если бы ему прислали оптический инструмент,
сконструированный так небрежно, как глаз, он отослал бы его назад изготовителю.
Однако «физиологические» недостатки глаза как фокусирующего устройства в
значительной мере компенсируются нейронными механизмами–контрастирования.
Астигматизм. Поверхность роговицы не симметрична относительно
оптической оси: обычно кривизна в вертикальной плоскости несколько больше, чем
в горизонтальной. Как следует из уравнения (1), это приводит к зависимости
преломляющей силы от угла падения лучей (астигматизм,
или астигмия). Если разница не
превышает 0,5 дп, такой астигматизм называют «физиологическим».
Сферическая аберрация. Как и у всех простых линз, у роговицы и хрусталика
фокусное расстояние больше в центральной части, через которую проходит
оптическая ось, и меньше на периферии. Возникающая за счет этого сферическая аберрация
делает изображение нерезким. Чем меньше диаметр
зрачка, тем меньше участие периферических частей диоптрической системы в
построении изображения, а следовательно, и искажения, вызванные сферической
аберрацией.
Хроматическая аберрация и аккомодация. Как и все простые линзы, диоптрический аппарат глаза преломляет
свет тем сильнее, чем короче длина его волны. Это приводит к хроматической аберрации. Для точной
фокусировки красных частей объекта требуется большая аккомодация, чем для
синих. Именно поэтому синие предметы кажутся более удаленными, чем красные,
находящиеся на том же самом расстоянии от наблюдателя. Архитекторы готических
храмов часто использовали эту физиологическую иллюзию при создании цветных
витражей: фон делался синим, а фигуры окрашивались в другие цвета, и как бы
выступали на передний план.
Рассеяние света и помутнение
диоптрического аппарата. В состав хрусталика и стекловидного
тела входят структурные белки и другие макромолекулярные вещества в коллоидном
растворе. Поэтому в диоптрическом аппарате происходит некоторое рассеяние света. Однако оно влияет на
зрительное восприятие только в случае очень ярких стимулов. В стекловидном теле
даже вполне здорового глаза есть зоны
помутнения, которые при рассматривании белого фона заметны как мелкие
сероватые кружки или пятна неправильной формы. Когда глаза движутся, они кажутся
пролетающими на светлом фоне мошками. У пожилых людей содержание воды в
хрусталике может снизиться до такой степени, что он, уплотнившись, станет
непрозрачным (старческая катаракта).
Если такому больному удалить хрусталик, он будет нормально видеть при
соответствующем подборе очков (примерно +13 дп для дали).
Миопия. Суммарная преломляющая сила диоптрического аппарата
здорового неаккомодированного глаза составляет 58,6 дп. При этом для получения
четкого изображения очень далекого объекта нужно, чтобы расстояние между
полюсом роговицы и центральной ямкой составляло 24,4 мм. Если осевая длина
глазного яблока больше, удаленные объекты невозможно точно сфокусировать,
поскольку фокальная плоскость находится перед
центральной ямкой (близорукость,
или миопия). Чтобы хорошо видеть
вдали, близоруким людям нужны очки с вогнутыми линзами (рис. 11.9).
Гиперметропия. Если при обычной преломляющей силе диоптрического
аппарата глаза его осевая длина слишком мала, возникает дальнозоркость (гиперопия, или гиперметропия). Лица с таким недостатком отчетливо видят удаленные
предметы, пользуясь механизмом аккомодации на ближний план. Однако диапазон
аккомодации у них недостаточен для точной фокусировки на сетчатке изображения
близко расположенных объектов. Чтобы компенсировать этот недостаток, требуются
очки с выпуклыми линзами (рис. 11.10) [12, 23, 26].

Рис. 11.9.
Миопия (близорукость) и ее коррекция с помощью вогнутой линзы. Для наглядности
длина глазного яблока преувеличена («осевая миопия»)

Рис. 11.10. Гиперметропия
(дальнозоркость) и ее коррекция с помощью выпуклой линзы
Измерение аномалий лучепреломления и
выписка рецепта на очки
Преломляющую силу диоптрического аппарата можно
измерить либо объективными, либо субъективными методами. Первые
(ретиноскопия, инфракрасная техника) относятся к области офтальмологии. Здесь
же мы кратко рассмотрим лишь субъективные
процедуры.
Субъективное измерение лучепреломления осуществляется, например, с помощью специальных
таблиц. Пациент рассматривает их одним глазом с расстояния 6 м. В случае
подозрения на близорукость перед его глазом ставят вогнутые линзы со все более
отрицательными значениями преломляющей силы (в диоптриях), пока не будет
достигнута нормальная острота зрения. Преломляющая сила корректирующей линзы примерно
равна ошибке лучепреломления. Пациенты с гиперметропией, как правило, хорошо
видят удаленные предметы. Им подбирают линзы с выпуклой поверхностью
(«плюсовые»), пока не появится нечеткость изображения. По количеству их
диоптрий можно определить степень гиперметропии.
Очки. Корректирующая линза и глаз вместе образуют составную оптическую
систему, к которой применимо уравнение (4). Если вместо «минусовых» очков для
коррекции зрения носить роговичные
контактные линзы, член d в уравнении (4)
станет меньше. Это значит, что преломляющая сила таких линз может быть
несколько меньше, чем у очков. При подборе последних нужно тщательно следить за
совпадением оптических осей глаза и корректирующей линзы (если смотреть прямо
перед собой). Для этого необходимо измерять расстояние между зрачками (56–70
мм) и подбирать соответствующую оправу.
Размер изображения предмета на сетчатке зависит от преломляющей силы
корректирующих линз. Если для левого и правого глаза она не одинакова,
изображения на двух сетчатках будут различаться по величине, что может привести
к нарушению бинокулярного зрения. В
этом случае нужно найти компромисс между оптимальной коррекцией и нормальным
бинокулярным зрением. Разница в преломляющей силе двух линз очков не должна
превышать 3 дп.
Если у человека ярко выражен «регулярный» астигматизм, т.е. закономерная разница
в кривизне роговицы вдоль разных ее диаметров, его можно скорректировать цилиндрическими линзами. Если же
астигматизм «нерегулярный» (кривизна роговицы варьирует «бессистемно»), его
лучше компенсировать контактными линзами.
Исследование внутренней структуры глаза
с помощью офтальмоскопа
Если в свет фар автомобиля попадет смотрящее на
водителя животное, его глаза могут показаться «горящими» за счет отражения
света от глазного дна (внутренней поверхности глазного яблока). Эта отражающая
способность глаза используется в офтальмоскопии.
На рис. 11.11 показана упрощенная схема хода падающих и отраженных лучей при
таком обследовании. Чтобы отчетливо видеть сетчатку пациента, врачу необходимо
настроить свои глаза, как на рассматривание удаленных предметов. Ошибки
лучепреломления глаз врача и пациента корректируют соответствующей оптикой.
На рис. 11.12 показан пример изображения, видимого при таких условиях. Диск зрительного нерва (papilla), сосуды
сетчатки и ее внутренняя поверхность увеличены примерно в 15 раз, поскольку
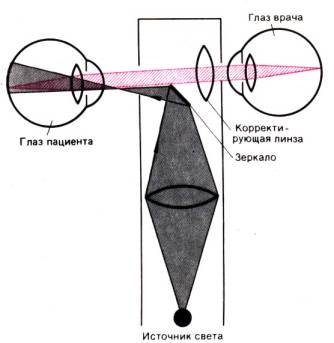
Рис. 11.11.
Сильно упрощенная схема метода прямой офтальмоскопии (вид сверху)
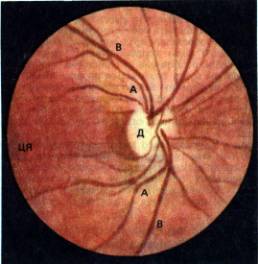
Рис. 11.12.
Глазное дно (часть сетчатки правого глаза) при обследовании методом прямой
офтальмоскопии. А–ветви центральной артерии сетчатки, В–ветви центральных вен
сетчатки, Д–диск зрительного нерва, ЦЯ–центральная ямка (W. Leydhecker. GrundriK der
Augenheilkunde, 20th Ed. 1979)
диоптрический аппарат действует как увеличительная
линза; изображение в данном случае прямое.
Чтобы получить
перевернутое изображение сетчатки, врач использует «непрямой» офтальмоскоп,
помещаемый на расстоянии около 80 см от исследуемого глаза. Параллельные лучи,
отраженные им, фокусируются выпуклой линзой с оптической силой от +13 до +15дп,
давая перевернутое действительное изображение сетчатки, на которое врач
аккомодирует свои глаза. При таком способе достигается
всего четырехкратное увеличение, однако можно увидеть более обширную часть
глазного дна, включая его периферические зоны.
Внутриглазное давление
Внешняя форма глаза и относительное расположение
отдельных частей его диоптрического аппарата почти неизменны из–за наличия
жесткой склеры и постоянства внутриглазного давления.
Секреция водянистой влаги. Давление внутри глазного яблока зависит в основном от
количества непрерывно вырабатываемой и отводимой из глаза водянистой влаги. За
счет ультрафильтрации (см. разд. 30.3) жидкость плазмы из
кровеносных капилляров цилиарного тела переходит в его межклеточное
пространство (рис. 11.5). Затем эпителиальные клетки цилиарного тела секретируют ее в виде водянистой влаги
в заднюю камеру глаза.
Оттуда водянистая влага перетекает в переднюю камеру,
а затем в венозную систему через трабекулярную сеть, находящуюся на краю камеры
(в месте соединения радужной оболочки и роговицы), и шлеммов канал.
Внутриглазное давление остается постоянным, если количества водянистой влаги,
выводимой из шлеммова канала и образующейся в цилиарном теле за тот же отрезок
времени, точно соответствуют друг другу. Если скорость образования водянистой
влаги нормальная, а ее отток затруднен, внутриглазное давление повышается. Его
патологическое повышение называется
глаукомой. При хронической (простой) глаукоме механически наиболее слабая
часть стенки глаза – решетчатая пластина–выпячивается наружу, ухудшая кровоснабжение волокон зрительного нерва. В случае
приступа острой глаукомы (глаукомы
«суженного угла») блокируется радужно–роговичный угол, резко повышается
внутриглазное давление и нарушается кровоток в сетчатке. Это ведет к ее либо
временному, либо необратимому поражению (слепоте).
Упругие силы, развиваемые радужкой, передаются на
радужно–роговичный угол (рис. 11.5) таким образом, что при напряженной радужке (суженном
зрачке) трабекулярная сеть и шлеммов канал расширяются. Поэтому лекарства,
вызывающие сужение зрачков, ускоряют отток водянистой влаги, а расширяющие их
(например, атропин)–замедляют. Таким образом, при подозрении на глаукому
следует всячески избегать расширяющих зрачок препаратов.
Тонометрия. Внутриглазное давление можно определить косвенным
способом, измеряя степень прогибания роговицы под действием стерженька с
определенными весом и диаметром
(импрессионная тонометрия) или же силу, которую нужно приложить, чтобы
сделать плоским небольшой участок роговицы
(аппликационная тонометрия). Если при нескольких повторных измерениях
внутриглазное давление превышает 20 мм. рт. ст. (2,66 кПа), оно считается
патологически повышенным. Во время приступа острой глаукомы оно может превышать
60 мм рт. ст. (8 кПа) [30, 37].
11.3.
Восприятие и обработка сигналов сетчаткой
В ходе эмбрионального развития сетчатка формируется за
счет выпячивания основания промежуточного мозга. Значит, это часть головного мозга.

Рис. 11.13.
Структура сетчатки приматов (по Boycott, Cowling. Proc. Roy. Soc. (Lond.), 1966, 166,
80, схематизировано) и схема ответов одиночных нейронов сетчатки на световой
стимул (Grusser, Fortschr. Ophthalmol., 1983, 80, 502).
НПМ–наружная пограничная мембрана, ВПМ внутренняя пограничная мембрана,
АЗН–аксоны зрительного нерва. Горизонтальные клетки образуют дополнительные
контакты с биполярами, не показанные на схеме
У позвоночных слой рецепторных клеток сетчатки
(палочек и колбочек) расположен на стороне, удаленной от стекловидного тела, и
находится в тесном механическом и функциональном контакте с клетками пигментного эпителия. Те в свою очередь прилегают к
сосудистой оболочке, так что служат важным путем для метаболитов рецепторных
клеток. Последние отделены от стекловидного тела слоями горизонтальных,
биполярных, амакриновых и ганглиозных клеток (рис. 11.13). Механически наиболее слабая часть сетчатки–граница между пигментным
эпителием и наружными сегментами фоторецепторов, где она может легко
«отслоиться». В отслоенной зоне наружные
сегменты рецепторов дегенерируют, и световосприятие здесь становится
невозможным, но если вовремя предприняты необходимые терапевтические меры,
восстанавливающие их постоянный контакт с пигментным эпителием, эти сегменты регенерируют и зрение также
восстанавливается.
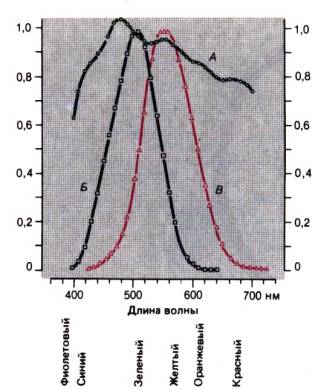
Теория двойственности зрения
Подстройку к сильно варьирующему уровню внешнего
освещения облегчает наличие двух систем сетчаточных рецепторов с разными
абсолютными порогами (теория двойственности).
В сумерках и ночью работают палочки
(скотопическое зрение), а при нормальном дневном свете–колбочки (фотопическое зрение). В первом случае цвета не различаются, хотя и при свете звезд
предметы не одинаковы по яркости. При фотопическом зрении различимы как их
яркость, так и окраска. Кривая спектральной чувствительности глаза имеет
максимум около 500 нм при скотопическом зрении, и около 550 нм при фотопическом
(рис. 11.4). Переход между скотопическим и фотопическим называют мезопическим
зрением. При нем возможно ограниченное цветоразличение [12].
Процесс трансдукции при зрении
Структура фоторецепторов. У человека слой рецепторов сетчатки состоит примерно
из 120 млн. палочек и 6 млн. колбочек (рис. 11.13). Плотность колбочек (их
число на единицу площади) максимальна в середине центральной ямки, а
палочек–вокруг этой ямки; в ней самой они полностью отсутствуют. Оба типа
рецепторов сходны по структуре в том смысле, что и у тех, и у других есть
наружный сегмент, состоящий примерно из тысячи мембранных дисков (палочки) или
мембранных складок (колбочки). Он соединяется с остальной частью клетки узкой
«ресничкой» (рис. 11.14, А, Б). На
поперечном срезе сетчатки наружные сегменты фоторецепторов образуют правильную
мозаику. В середине центральной ямки диаметр наружного сегмента колбочки
составляет примерно 2 мкм, что соответствует углу зрения около 0,4'. К
периферии сетчатки этот диаметр увеличивается.
Зрительные пигменты.
Молекулы зрительных пигментов очень упорядочение включены в двойной липидный
слой мембранных дисков наружных сегментов (рис. 11.14). У приготовленного в
темноте раствора зрительного пигмента палочек (родопсина, или «зрительного
пурпура») красный цвет, так как он особенно сильно поглощает зеленые и синие
лучи. Более точно можно оценить эту способность, сняв спектр поглощения зрительного пигмента. У родопсина два его
максимума–один в видимой части спектра (около 500 нм), другой–в
ультрафиолетовой (около 350 нм).
Кривые поглощения зрительных пигментов одиночных
фоторецепторов получают с помощью микроспектрофотометрии:
иссекают кусочек сетчатки и под микроскопом направляют очень узкий пучок света
с разной длиной волны сквозь изолированный наружный сегмент рецептора на
высокочувствительный фотоэлемент (рис. 11.15). Этот способ показал, что:
1) у зрительных пигментов палочек и колбочек разные
спектры поглощения;
2) спектр поглощения палочек такой же, как у
родопсина, и достаточно близок к кривой спектральной чувствительности
скотопического зрения (рис. 11.4). Родопсин состоит из гликопротеина (опсина) и
хромофорной группы – 11 –цис–ретиналя, т. е. альдегида витамина А (ретинола);
3) есть три
типа колбочек, различающихся своими зрительными пигментами (рис. 11.15).
Выцветание и регенерация зрительных
пигментов в результате поглощения света. Процесс трансдукции в фоторецепторе начинается с поглощения фотона
π–электронами сопряженных двойных связей ретиналя. В результате молекула
переходит на более высокий энергетический уровень и испытывает более сильные
колебания. При этом с вероятностью 0,5–0,65 («квантовая эффективность»)
происходит стереоизомеризация ретиналя,
т.е. его переход из цис- в полностью транс–форму. Затем вся молекула пигмента
в несколько этапов разрывается с образованием в конечном счете ретинола и опсина (см. учебники
биохимии). Чтобы использовать данный первичный фотохимический процесс для
передачи сигналов, он должен быть «превращен» в изменение

Рис. 11.14.
Схема строения палочки (А) и колбочки
(б) в сетчатке позвоночных. Структура мембранного диска наружных сегментов
палочек и мембранных складок наружных сегментов колбочек показана в увеличенном
виде. В. Структура диска или
плазматической мембраны фоторецептора. Ml–метародопсин I; МII–метародопсин II; Р–родопсин; М–внутриклеточный
медиатор
мембранного потенциала клетки. Детали превращения еще
не вполне ясны. Согласно одной из гипотез, при нем «активируются» связанные в
мембранных дисках ионы кальция, которые в результате диффундируют из них в
плазматическую мембрану наружного сегмента палочки или же–в случае колбочек–из
одного участка плазматической мембраны в другой (рис. 11.14,В). В итоге
проводимость мембраны для мелких ионов, особенно натрия, уменьшается и возникает
вторичный рецепторный потенциал, обсуждаемый ниже.
Более поздняя гипотеза основана на представлении о
сравнительно высокой натриевой проводимости мембраны наружного сегмента в
темноте и, следовательно, прохождении через нее «темнового тока». Эта
проводимость зависит от взаимодействия циклического 3'–5'–гуанидин–монофосфата
(цГМФ) с белковыми молекулами
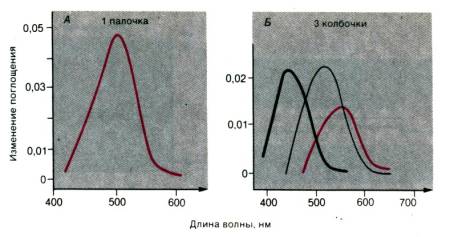
Рис. 11.15. Результаты
микроспектрофотометрических измерений спектров поглощения одиночных рецепторов
сетчатки человека (образцы взяты во время операций). Кривые–дифференциальные
спектры (разница спектров поглощения до и после выцветания). Можно различить
три типа колбочек (по Brown, Wald, Science, 1964,
144, 45, схематизировано)
натриевых каналов мембраны рецептора; цГМФ заставляет
их оставаться «открытыми». Тогда первичный фотохимический процесс состоит в
восстановлении за несколько миллисекунд цГМФ за счет цепочки ферментативных
реакций: натриевая проводимость уменьшается и возникает гиперполяризационный
вторичный рецепторный потенциал. Предполагается, что при. этом молекула
родопсина, превратившись в метародопсин II за счет поглощения фотона, в крайне
быстрой последовательности активирует множество молекул фермента
(«трансдуцина»), что приводит к начальному «усилению» первичного
фотохимического процесса, поскольку «трансдуцин» управляет цГМФ [4, 5, 8, 34,
46, 53, 56а]. Первичный рецепторный
потенциал фоторецепторов и фотохимические компоненты световой и темновой
адаптации. Конформационное изменение молекулы родопсина генерирует с очень
коротким латентным периодом (меньше 1 мс) электрический потенциал, называемый первичным (ранним)
рецепторным потенциалом (РРП) (рис. 11.16). В нем несколько
компонентов, которые можно выявить, постепенно снижая температуру сетчатки.
Когда зрительный пигмент распадается на ретинол и опсин, происходит его
ресинтез за счет цепи поглощающих энергию ферментативных реакций. Если на сетчатку
падает постоянный поток фотонов, устанавливается динамическое равновесие между процессами выцветания зрительного
пигмента под действием света и его регенерации с участием ферментов. Если
сетчатки достигает меньше света, оно сдвигается в сторону более высокой
концентрации родопсина. Это фотохимическая
основа хорошо всем известной темновой
адаптации глаза. В ходе нее амплитуда РРП, вызываемого световыми стимулами
постоянной интенсивности, повышается, поскольку возрастает вероятность
соударения падающих фотонов с молекулами невыцветшего родопсина [8].

Рис. 11.16.
Ранний рецепторный потенциал фоторецепторов бурундука, записанный при двух
различных температурах (по Pak, Ebrey: J. gen. Physiol., 1966,
49, с изменениями). Амплитуда различных его компонентов возрастает примерно
пропорционально логарифму силы световой вспышки. Стрелкой показан момент
вспышки
Вторичный рецепторный потенциал. Если РРП генерируется
за счет синхронных конформационных изменений молекул зрительного пигмента, то вторичный (поздний) рецепторный
потенциал ПРП, не связанный с РРП непосредственно, это изменение
мембранного потенциала фоторецептора. В темноте он составляет от —25 до —40 мВ; под действием света происходит гиперполяризация. Амплитуда этого
рецепторного потенциала растет с увеличением интенсивности светового стимула
(рис. 11.17), причем у палочек медленнее, чем у колбочек. Другими словами,
временная инерция палочковой системы больше, чем у колбочковой. Неодинаковая
спектральная чувствительность вторичных рецепторных потенциалов различных типов
колбочек (рис. 11.18) подтверждает результаты микро–спектрофотометрии и говорит
в пользу трихроматической
(трехкомпонентной) теории цветового зрения, т.е. присутствия трех типов
колбочек с разными спектральными чувствительностями. Спектральная
чувствительность рецепторных потенциалов палочек примерно соответствует спектру
поглощения родопсина с максимумом около 500 нм. У позвоночных рецепторный
потенциал фоторецепторов в отличие от других рецепторов связан

Рис. 11.17.
Поздний рецепторный потенциал фоторецепторов сетчатки позвоночных. А. Внутриклеточная регистрация
рецепторных потенциалов колбочки и палочки (схематично). 6. Рецепторный
потенциал колбочки сетчатки черепахи в ответ на световые вспышки длительностью
10 мс с возрастающей интенсивностью (относительные интенсивности: а = 1, б = 4,
в = 16). В. Кривая амплитуды
рецепторного потенциала одиночной колбочки сетчатки черепахи. Относительная
амплитуда (ось ординат) увеличивается
пропорционально десятичному логарифму относительной интенсивности стимула, Ig (ось абсцисс)
в сравнительно узком диапазоне. Показана также схематическая кривая зависимости
величины возбуждения ганглиозной клетки сетчатки с оп–центром от силы света (Б, В: по Baylor, Fuortes
J. Physiol. (Lond.) 1970, 207)
не с деполяризацией, а с гиперполяризацией мембраны в
ответ на адекватную стимуляцию (свет).
Амплитуда (А) вторичного рецепторного потенциала
коррелирует с интенсивностью светового стимула (Is) (т.е. с числом фотонов, падающих в единицу времени
на единицу площади) следующим образом (рис. 11–17, Б, В):
A= α Is /1+k Is =[мв] (9)
Эта гиперболическая функция («закон Геринга») в
диапазоне промежуточных интенсивностей приближается к логарифмической, которая
в общей сенсорной физиологии известна как
закон Вебера–Фехнера :
A=k*–log Is / Io [MB], (10)
где Io пороговая интенсивность стимула, зависящая от уровня адаптации. Константы a, k и k* в уравнениях (9) и (10) меняются вместе с длиной волны
монохроматического света. Эта зависимость примерно соответствует кривым
спектральной
чувствительности рецепторных потенциалов (рис. 11.18)
[8, 36].
Роговично–сетчаточный потенциал и
электроретинограмма (ЭРГ)
С помощью макроэлектродов от всего глаза можно
отводить два функционально различных типа электрических потенциалов: постоянный – между роговицей и более
отрицательной сетчаткой–и
электроретинограмму (ЭРГ). Первый обусловлен главным образом разностью
потенциалов между склеральной стороной клеток пигментного эпителия и
внутренними сегментами фоторецепторов. Другими словами, он отражает суммарный
электрический ток, проходящий через плазматические мембраны пигментных клеток и
фоторецепторов, поэтому меняется в зависимости от уровня адаптации сетчатки.
ЭРГ–это колебания отводимого от всего
глаза напряжения, вызываемые увеличением или уменьшением количества падающего
на сетчатку света. ЭРГ включает несколько «волн»: а, b, с и d (рис.
11.19). Волна а возникает, по–видимому, за счет суммации рецепторных
потенциалов; более медленная b–волна отражает
изменения, главным

Рис. 11.18.
Спектральная чувствительность рецепторных потенциалов сетчатки рыбы. Показаны
средние значения (со стандартным отклонением) результатов для трех разных типов
колбочек (по [61] с изменениями)

Рис.
11.19. Электроретинограммы сетчатки
человека (no Hanitzsch et al.. Vision Res. 1966, 6,
245, схематизировано)
образом мембранных потенциалов биполярных и глиальных
(мюллеровых) клеток, с–волна – мембранных потенциалов клеток пигментного
эпителия при «включении света» (оn–эффект), а
d–волна мембранных потенциалов фоторецепторов и биполярных клеток при
«выключении света» (оff–эффект). Если
длительность светового стимула превышает 0,3 с, с–волна появляется еще в ходе
его действия и off–эффект накладывается на нее. При коротких световых вспышках b– и d–волны совпадают [8, 10].
ЭРГ обращаемого узора регистрируется при рассматривании узора, светлые и
темные участки которого периодически меняют яркость на противоположную
(например, при обращении цвета светлых и темных шахматных полей на телеэкране).
Средняя яркость такого стимула неизменна, поэтому ЭРГ в данном случае
генерируется в основном ганглиозными клетками сетчатки, т. е. нейронными
элементами, особенно сильно реагирующими на изменения
контраста и контрастные границы.
Если ганглиозные клетки дегенерируют, ЭРГ обращаемого узора исчезает. Такой
симптом в медицине обычно связан со слепотой или с резким ухудшением остроты
зрения.
Рецептивные поля нейронов сетчатки
Нейронная сеть сетчатки. Вторичный рецепторный потенциал передается через
синаптические контакты от рецепторов биполярным и горизонтальным клеткам с
помощью химического медиатора.
Рецептивные поля (РП) последних распространяются на несколько
фоторецепторов. Если осветить фоторецепторы
центра РП биполярных клеток оn–типа, мембрана этих биполяров деполяризуется (рис. 11.13), если периферии РП–гиперполяризуется.
Гиперполяризация осуществляется косвенно–за счет контактов между
горизонтальными и биполярными клетками или же через петлю обратной связи между
горизонтальными клетками и синаптическими пузырьками колбочек.
РП горизонтальных клеток, как правило, сравнительно
велики. Функционально они связаны друг с другом. Некоторые горизонтальные
клетки (L–muпа)
гиперполяризуются при попадании света на их РП независимо от его спектрального
состава; другие при освещении либо деполяризуются, либо гиперполяризуются в
зависимости от того, какой части спектра соответствует стимул (цветоспецифичные
красно–зеленые или сине-желтые горизонтальные клетки).
Биполярные клетки off–типа гиперполяризуются при попадании светового стимула в
центр их РП и деполяризуются, когда он падает на периферию РП (рис. 11.13).
Сетчатка содержит примерно равные количества on– и ой–биполяров. Функциональная организация их слоя
определяется двумя важнейшими принципами обработки зрительной информации
нейронами.
1. Возбуждение фоторецепторов фотонами передается в
ЦНС по двум независимым нейронным «каналам»: через оп–систему, активируемую,
когда световая стимуляция в центре РП сильнее, чем на периферии или чем
предыдущая стимуляция центра, и через off–систему, активируемую при уменьшении
освещения.
2. Рецептивные поля многих биполяров организованы антагонистически, т.е. световой стимул
вызывает их диаметрально противоположную
реакцию в зависимости от того, попадает он на периферию или в центр РП. Такая
пространственная организация РП обеспечивается двумя способами: конвергенцией
сигналов от нескольких рецепторов на одной биполярной клетке и опосредованными
тормозными сигналами с периферии РП, доходящими до биполяров через
горизонтальные клетки (латеральное торможение).
Этот принцип
конвергенции сигналов и латерального торможения действует и на следующем
уровне обработки информации в сетчатке (ганглиозные и амакриновые клетки, рис.
11.13). Как правило, несколько on– («прямое»
возбуждение) и off– («прямое» торможение)
биполяров конвергируют на одну ганглиозную клетку с оп–центром, однако
значительно больше биполярных клеток косвенно связаны с последней посредством
контакта с амакриновыми клетками формируя
периферию ее с РП (латеральные оп–торможение и оп–возбуждение). Ганглиозные
клетки с off–центром возбуждаются off–биполярами, прямо тормозятся оn–биполярами (центр РП), «латерально» тормозятся
off–амакриновыми клетками и, наконец, латерально возбуждаются амакриновыми
клетками, возбуждаемыми оn–биполярами
[11, 17, 18, 33, 49].
Классы ганглиозных клеток сетчатки
Стимуляция
ахроматическим светом сетчатки млекопитающих позволила обнаружить три
основных класса ганглиозных клеток. Два из них описаны выше—их рецептивные поля
организованы антагонистически.
Ганглиозные клетки с оn–центром деполяризуются
в ответ на освещение центра РП; эта деполяризация в аксонном холмике
преобразуется в последовательность потенциалов действия (по закону «все или
ничего») (рис. 11.13, 11.20). С другой стороны, как освещение периферии РА, так
и «выключение света» в центре РП вызывают
гиперполяризацию их мембранного потенциала и, следовательно, кратковременное
торможение нейронной активности. Если центр и периферия РП освещаются одновременно, доминирует реакция
центра. Однако при этом активация меньше, чем при освещении только центра РП,
поскольку его возбуждение и торможение периферии РП суммируются (рис. 11.20).
Рецептивные поля
ганглиозных клеток с off–центром функционально противоположны только что
описанным. Освещение их центра приводит к «прямому» торможению, а ослабление
падающего света– к «off–активации».
Освещение периферии РП ведет к латеральной активации, а «выключение света» на
периферии– к кратковременному «латеральному» торможению частоты импульсации
нейрона. При одновременном освещении центра и периферии эти эффекты
суммируются, причем торможение и off–активация, связанные с центром РП, как правило,
доминируют (рис. 11.20).
Ганглиозные клетки on–off–типа обычно дают короткий оn–ответ на стационарный световой стимул и короткую
off–реакцию на ослабление света. В этот класс клеток входят, например, нейроны,
чувствительные к движению, особенно хорошо воспринимающие перемещение через их
рецептивное поле границы света и темноты. При этом степень возбуждения зависит
от угловой скорости стимула.
Еще одна классификация ганглиозных клеток основана на скорости проведения сигналов по их аксонам.
У большинства крупных ганглиозных клеток со сравнительно толстыми
миелинизированными аксонами она высока. На освещение рецептивного поля они дают
короткий «фазический» ответ (нейроны
I класса латентности, или Y–нейроны).
Гораздо многочисленнее мелкие ганглиозные клетки с более тонкими
миелинизированными аксонами, которые на освещение центра РП реагируют «тоническим» возбуждением или
торможением (нейроны II класса латентности, или Х–нейроны). Среди Х– и Y–нейронов клетки с on– и off–центром одинаково часты. Наконец, в сетчатке
есть ганглиозные клетки с тонкими, лишь слегка миелинизированными аксонами (III класса латентности, или W–нейроны). К этому классу в
основном и относятся on–off–нейроны,
чувствительные к движению.
Даже если не рассматривать цветоспецифичные реакции
ганглиозных клеток сетчатки, очевидно, что она представляет собой сложную
нейронную систему. Уже внутри нее самой
изображение, активирующее входной слой рецепторов, преобразуется в несколько
типов возбуждения различных ганглиозных клеток [10, 49, 52].
11.4.
Нейрофизиология и психофизика восприятия света и темноты
Когда в ясный день на солнце набегает облако, мы
замечаем уменьшение освещенности, к которому вскоре адаптируемся. Даже при стократном изменении силы света
воспринимаемые относительные светлота и цвета окружающих объектов изменяются
весьма незначительно. В условиях фотопического зрения восприятие контраста и
цвета сравнительно независимо от средней освещенности. Другими словами, вид
предмета определяется отражательной способностью его поверхностей, а не
абсолютным количеством отраженного от них света. Данный механизм важен для
понимания удивительной неизменности видимого нами мира.
Если при
постоянном уровне адаптации освещается определенная область сетчатки,
наблюдается приблизительно логарифмическая зависимость между воспринимаемой субъективной светлотой светового пятна
и его измеренной яркостью (уравнения
9, 10). С помощью микроэлектродных записей показано, что такой же зависимостью
описывается частота импульсации нейронов с оn–центрами (рис. 11.17, 5). У нейронов с off–центрами она также приблизительно соответствует
логарифмической функции предшествующего снижения стимуляции при «выключении
света». В целом отмечается линейная
зависимость между субъективной

Рис. 11.20.
Функциональная организация рецептивных полей ганглиозных клеток сетчатки
млекопитающих. При анализе рецептивных полей небольшие пятна света (показаны белым) проецировались либо на центр,
либо на периферию РП. Световые стимулы вызывают разный ответ у нейронов с оn– и off–центром. Когда обе части РП освещены
одновременно, возбуждающий и тормозный процессы, связанные с освещением центра
и периферии, суммируются. Однако преобладает ответ, вызываемый стимуляцией
центра РП
светлотой и средней частотой импульсации
нейронов оn–системы и
между субъективной темнотой и средней частотой импульсации нейронов off–системы.
Зная это простое правило корреляции нейронной
активности и восприятия, можно объяснить целый ряд элементарных зрительных
феноменов.
Нейрофизиологическая основа
одновременного контраста
Рассмотрим первый пример корреляции между зрительным
восприятием и возбуждением on– и
off–нейронов. Серое пятно на белом фоне кажется темнее, чем на черном (рис.
11.21). Вдоль границы между темным и светлым наблюдаются полосы Маха
(пограничный контраст): ближайшая к ней светлая часть кажется светлее, а
ближайшая темная–темнее, чем более удаленные от границы. Исходя из
функциональной организации РП ганглиозных клеток сетчатки, можно предложить
следующее объяснение этого простого примера одновременного контраста (рис.
11.22). Одновременный контраст–важный механизм, обеспечивающий частичную
функциональную компенсацию физиологических недостатков диоптрического аппарата,
описанных на, и в результате повышающий остроту зрения [35].
Изменение остроты зрения и организации
рецептивных полей при изменениях окружающей освещенности
Каждый знает по опыту, что острота зрения снижается от
центра поля зрения к его периферии и что она гораздо хуже при скотопическом зрении,
чем при фотопическом. В фотопических
условиях острота зрения снижается при уменьшении средней освещенности.
Вы
сможете прочесть такой мелкий шрифт, только если будете смотреть прямо на него
.
Зависимость контрастности восприятия и остроты зрения
от средней освещенности можно объяснить реорганизацией РП ганглиозных клеток
сетчатки (рис. 11.23). Очевидно, что порог прямого возбуждения (за счет центра
РП) соответствует меньшей силе света, чем для латерального торможения, однако
эффективность последнего при росте освещенности увеличивается быстрее, чем у
возбуждения. Если пространственная организация РП соответствует суммации
возбуждения и торможения (рис. 11.23), диаметр функционального центра РП
уменьшается при возрастании средней освещенности. Далее, исходя из соотношения
числа рецепторов и ганглиозных клеток (в среднем примерно 125:1), очевидна
весьма выраженная конвергенция сигналов
из слоя рецепторов в слое ганглиозных клеток. Однако в зоне центральной ямки
она значительно меньше. Соотношение конвергенции и латерального торможения в
этой области сетчатки при оптимальных условиях освещения позволяет достичь
остроты зрения, близкой к теоретическому пределу,

Рис. 11.21.
Рисунки, иллюстрирующие явление одновременного яркостного контраста
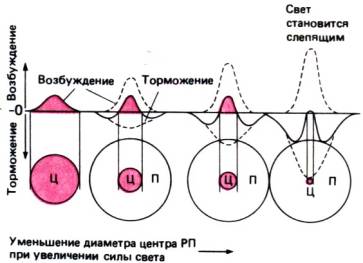
Рис. 11.23.
Пространственная организация рецептивного поля ганглиозной клетки оn–типа, объясняемая суперпозицией пространственных
распределений возбуждающего и тормозного процессов. У возбуждения порог ниже,
чем у торможения. По мере роста освещенности в надпороговой области, если
торможение увеличивается быстрее возбуждения, центр РП будет функционально
уменьшаться. При очень ярком (слепящем) свете преобладает торможение: световая
реакция ганглиозной клетки оп–типа тормозится и зрительное восприятие формы
нарушается. При оптимальном уровне световой адаптации размеры центра РП
минимальны, следовательно, острота зрения максимальна (Grusser, Fortschr. Ophthalmol., 1983, 80, 502). Ц–центр РП; П–периферия РП

Рис.11.22.
Схема механизма одновременного контраста. А–В.
Возбуждение нейрона с on–центром в
зависимости от положения в РП границы темного и светлого. Максимальная
активация соответствует случаю Б,
когда граница между темным и светлым совпадает с границей между центром и
периферией РП. Числа соответствуют относительной амплитуде возбуждения (без
учета спонтанной активности нейрона). Г. Зависимость
возбуждения нейронов зрительной системы от положения в РП границы темного и
светлого (ось абсцисс). Эти
упрощенные кривые отражают ответы нейронов сетчатки с on– и off–центрами, а также «контрастовых» нейронов ЛКТ
с оn–центрами
определяющемуся пространственной мозаикой популяции
колбочек –0,5', что примерно соответствует диаметру наружных сегментов
находящихся здесь фоторецепторов.
Снижение остроты зрения–не единственное отрицательное
последствие неадекватного освещения читаемой книги или рабочего места. Из–за кажущейся нерезкости изображения
рефлекторно включаются механизмы аккомодации, которые, разумеется, оказываются
в данном случае неэффективными. Когда система аккомодации непрерывно напряжена,
легко может возникнуть головная боль. Поэтому важно обеспечить надлежащее освещение рабочего места и избегать при этом
источников света с неустойчивой яркостью.
Световая и темновая адаптация,
послеобразы влияние сверхъярких стимулов
Когда средняя освещенность изменяется,
чувствительность зрительной системы приспосабливается к новым условиям. Когда человек выходит из
ярко освещенной комнаты на темную улицу, в первый момент он не способен
различать окружающие предметы, однако через некоторое время становятся видимыми
по крайней мере их очертания.
Во время
темновой адаптации абсолютная чувствительность зрительной системы медленно
возрастает, одна ко острота зрения всегда становится значительно хуже чем при
дневном свете. Определить временной ход такой адаптации можно, периодически
измеряя пороговые значения силы света (рис. 11.24). Низкая скорость адаптации
хорошо согласуется с постепенным снижением освещенности среды при наступлении
сумерек. В ходе темновой адаптации палочковая система становится намного
чувствительнее колбочковой. После долгого пребывания

Рис. 11.24.
Кривые темновой адаптации человека. А. Усредненная
кривая для девяти здоровых испытуемых. Б.
Кривая при полной цветовой слепоте, полученная для области сетчатки на 8° выше
центральной ямки. В. Кривая
колбочковой системы человека с нормальным цветовым зрением (красный световой
стимул в области центральной ямки). Для кривой (Б) ось времени (абсцисс)
надо сдвинуть вправо на 2 мин (А, Б:
по данным Е. Auerbach, Лаборатория исследования
зрения, Иерусалим, 1973)
в полной темноте (более 45 мин) абсолютный порог зрительной чувствительности достигает уровня 1–4
фотонов в секунду на фоторецептор. В скотопических условиях слабые световые
стимулы лучше различаются не центральной ямкой, а окружающей ее частью
сетчатки. Поэтому тусклые звезды видны только тогда, когда их изображения
проецируются на эту область; если смотреть прямо на них, они «исчезают».
У животных, хорошо приспособленных к ночному образу
жизни, чувствительность адаптированной к темноте сетчатки усиливается в 5–20
раз за счет слоя отражающих клеток (tapetum lucidum),
расположенного между пигментным эпителием и сосудистой оболочкой склеры. Этот
слой отражает фотоны, проходящие мимо рецепторов, и возвращает их к последним.
Его пигмент желтовато–зеленый, поэтому глаза кошек и собак «светятся» именно
таким цветом.
Процесс, обратный описанному, называется световой адаптацией; он протекает
значительно быстрее. Когда адаптированный к темноте человек входит в ярко освещенную
комнату, его зрительная система приспосабливается к новым условиям освещения за
несколько секунд. Если разница в освещенности слишком велика, может наступить
временное ослепление с ухудшением
восприятия формы [8, 13, 16, 25].
Механизмы световой и темновой адаптации.
Помимо сдвига равновесия между выцветшим и невыцветшим
пигментом фоторецепторов процессы световой и темновой адаптации включают важные
нейронные механизмы. Зрение «переключается» с колбочковой системы на палочковую
с помощью горизонтальных клеток. Кроме того, во время темновой адаптации
увеличивается функциональный размер центров РП сетчатковых нейронов (рис.
11.23). Еще один нейронный компонент световой и темновой адаптации –
зависимость размера зрачка от
средней освещенности .
Локальная адаптация и послеобразы. Локальная адаптация соответствует случаю, когда при
постоянной средней освещенности
среды в разных частях сетчатки она неодинакова. Если центр фигуры, изображенной
на рис. 11.25, фиксировать в течение примерно 30 с, то, переводя взгляд на
белый или серый фон, можно в течение нескольких секунд наблюдать негативный послеобраз. То, что было на
исходной фигуре темным, кажется светлым, и наоборот. Участки сетчатки, на
которые во время фиксации попадают темные фрагменты изображения, становятся
чувствительнее соседних, воспринимавших его светлые детали.
Послеобразы сохраняются довольно долго, если
ограниченная зона сетчатки освещалась очень сильно или достаточно долго.
Локальная адаптация к цветным узорам
приводит к появлению послеобразов, окрашенных в дополнительные цвета :
«Когда я вернулся в гостиницу поужинать, в мою комнату
вошла и встала недалеко от меня полная горничная с ослепительно белым лицом,
черными волосами и в алом платье. Я внимательно смотрел на нее в наступающих
сумерках. Затем, когда она вышла, я увидел на белой стене против меня черное
лицо, окруженное светлым ореолом, а одежда этой совершенно новой фигуры
казалась прекрасного сине–зеленого цвета». (Goethe, Zur Farbenlehre, I,
52.)
После воздействия яркой вспышки света можно наблюдать
быструю последовательность позитивных (светлых) периодических послеобразов, разделенных негативными темными
интервалами. Особенно легко. наблюдать их чередование, следя за узкой
движущейся полоской света. Возбуждение ганглиозных клеток сетчатки с on– и off–центрами хорошо коррелирует с периодичностью
этих послеобразов (рис. 11.26). Периодичность возбуждения частично обусловлена
обратной связью в пределах сетчатки за счет
интерплексиформиых клеток (рис. 11.13) [51].
Воздействие сверхъярких стимулов. При внезапном сильном засвете сетчатки (например,
ночью, фарами движущегося навстречу автомобиля) может возникнуть позитивный
послеобраз такой интенсивности, что зрительное восприятие формы временно
нарушится. При таком внезапном ослеплении срабатывает рефлекс смыкания век, дуга которого проходит через нервные
соединения сетчатки с подкорковыми зрительными центрами и нейронами лицевого
ядра. Сильное ослепление увеличивает секрецию слезной жидкости .
Фосфены давления
Если в полной темноте деформировать глазное яблоко,
нажимая пальцем на его край, сначала возникнет ощущение света в части поля
зрения, противоположной деформированной стороне. Если продолжать надавливать на

Рис.11.25. Рисунок,
демонстрирующий возникновение послеобраза. Если в течение примерно 30 с
фиксировать центр геометрической .фигуры справа, а затем перевести взгляд в
центр окружности слева, можно увидеть негативный послеобраз правой фигуры

Рис. 11.26.
Схема, иллюстрирующая корреляцию между фазическими послеобразами после коротких
световых вспышек и нейронной активностью ганглиозных клеток сетчатки. Нейроны с
оn–центрами (оn–Х и on–Y) возбуждаются
при восприятии первоначальных образов и светлых послеобразов, периодически возникающих
после стимуляции. Клетки с off–центрами посылают импульсы во время темных фаз
между послеобразами (по [51] с изменениями)
глазное яблоко, свет постепенно распространится по
всей сетчатке, причем вместе с ним появятся движущиеся световые облака и неподвижные яркие точки. Это так называемый
монокулярный фосфен давления, вызываемый деформацией сетчатки. Наблюдение
его–один из старейших известных экспериментов в сенсорной физиологии. Он был
впервые описан еще до Сократа философом и врачом Алкмеоном из Кротона (V в. до
н.э.). По–видимому, такие фосфены возникают следующим образом. Деформация
глазного яблока растягивает горизонтальные клетки сетчатки, повышая натриевую
проводимость их мембраны, что ведет к их деполяризации. За счет контактов между
горизонтальными и биполярными клетками оn–биполяры деполяризуются, а off–биполяры
гиперполяризуются. В результате этого ганглиозные клетки оп–системы
активируются, а off–системы – тормозятся, В
соответствии с вышеупомянутым правилом
корреляции человек видит свет [49].
Временные особенности передачи сигналов
в сетчатке
В век фильмов, телевидения и дисплеев временные
аспекты зрительного восприятия приобретают особую роль, поскольку все эти
изобретения создают воспринимаемые образы путем быстрого чередования вспышек
света, т.е. за счет прерывистой стимуляции сетчатки. Частотой слияния мельканий (или критической частотой мельканий,
КЧМ) называют наименьшую частоту поступления световых стимулов, при которой
испытуемый уже не воспринимает их по отдельности, т.е. они сливаются. При скотопическом (палочковом) зрении КЧМ
равна 22–25 стимулов в секунду, а при фотопическом
повышается примерно пропорционально логарифму яркости, степени модуляции и
стимулируемой площади, достигая 80 стимулов в секунду. Частота слияния
мельканий для ганглиозных клеток сетчатки подчиняется тем же закономерностям,
что и субъективная КЧМ. Световые стимулы с частотой 5–15 Гц вызывают особенно
сильное возбуждение нейронов сетчатки и первичной зрительной коры. Поэтому
мелькающий свет может вызвать у некоторых больных эпилепсией судороги [10, 17,
18].
Многие современные источники света (люминесцентные
лампы, телевизионные экраны, мониторы компьютеров) испускают прерывистые
световые стимулы с частотой 50–120 Гц, которые могут вызвать неустойчивое
возбуждение сетчатки, если глаза наблюдателя одновременно движутся по источнику света. Особенно неблагоприятна
длительная работа с монитором компьютера;
в этом случае должно строго выполняться требование
появления изображения на экране с частотой выше 100 Гц. Дело в том, что при
смещении изображения на сетчатке во время саккад ее прерывистое освещение
вызывает кажущееся снижение остроты
зрения. Безуспешные попытки системы аккомодации глаз скорректировать этот
кажущийся дефект могут привести к головной боли у того, кто смотрит на экран
несколько часов подряд.
11.5.
Обработка сигналов в центральных отделах зрительной системы
Центральные зрительные пути
Зрительная информация передается в головной мозг по
аксонам ганглиозных клеток сетчатки, которые образуют зрительный нерв. У
человека последний состоит примерно из 1 млн. волокон–миелинизированных, слабо
миелинизированных и немиелинизированных аксонов. Правый и левый зрительные
нервы сливаются у основания черепа, образуя
зрительный перекрест (хиазму) (рис. 11,27). Здесь нервные волокна, идущие
от носовых половин сетчаток, пересекаются и переходят на противоположную
сторону головы, а волокна от височных половин сетчаток продолжаются
ипсилатерально, объединяясь с пересекшимся пучком аксонов

Рис. 11.27.
Схема зрительных путей в головном мозгу человека. Справа показаны эфферентные связи между зрительной корой и
подкорковыми структурами. Области зрительной коры левого и правого полушарий
соединены друг с другом аксонами, проходящими через мозолистое тело (МТ).
Стрелки, отмеченные буквами, указывают на места возможных повреждений,
приводящих к выпадениям поля зрения, показанным на рис. 11.35
контралатерального зрительного нерва в зрительный тракт. Он ведет к первым
центральным станциям зрительного пути–
латеральным коленчатым телам, верхним холмикам четверохолмия, ядру зрительного
тракта (ЯЗТ), ядрам вспомогательного
зрительного тракта, претектальной области ствола мозга и гипоталамусу. Эти
связи выполняют следующие функции.
1. У человека наиболее важны и многочисленны связи
сетчатки с латеральным коленчатым телом,
которое состоит из двух
магноцеллюлярных (крупноклеточных) и четырех парвоцеллюлярных (мелкоклеточных) слоев. Непересекающиеся аксоны ипсилатерального зрительного нерва
оканчиваются в трех из этих слоев, а
пересекшиеся (контралатерального зрительного нерва) в трех других. Аксоны
большинства клеток коленчатого тела проходят через зрительную лучистость к нервным клеткам первичной зрительной коры (поле 17, или VI, затылочной коры
головного мозга). Та в свою очередь соединяется с вторичной зрительной корой (поле V2), третичной
зрительной корой (поля V3, V3a) и с интегративными зрительными зонами
затылочно–теменной и затылочно–височной коры головного мозга. Последние будут
описаны ниже. Проекция зрительного пути в
латеральном коленчатом теле участвует в распознавании объектов, цветовом
зрении, восприятии движения и в стереоскопическом восприятии глубины.
1. Связи между сетчаткой и гипоталамусом служат для сопряжения эндогенного циркадианного ритма сна I бодрствования
со сменой дня и ночи, а также для участия в управлении эндокринной системой. За
счет связей гипоталамуса с эпифизом сетчатка влияет также на пигментацию кожи.
3. Часть связей между сетчаткой и претектальной областью служит для регуляции диаметра зрачка.
4. Аксоны зрительного нерва оканчиваются на нервных
клетках в претектальных ядрах
«вспомогательного зрительного тракта». Те соединены с центрами ствола
мозга, управляющими взглядом, главным образом вертикальными и вергентными движениями глаз.
5. Проекция сетчатки в верхних холмиках четверохолмия служит для управления рефлекторными
саккадическими движениями глаз. Аксоны нейронов верхних холмиков идут в
глазодвигательные центры ствола мозга и в зрительную часть подушки. Зрительные
сигналы, проходящие по этим таламическим зонам, достигают в конечном счете теменных зрительных ассоциативных зон .
6. Ганглиозные клетки сетчатки, аксоны которых
оканчиваются на нейронах ядра
зрительного тракта (ЯЗТ), относятся преимущественно к специфично
реагирующим на движение (on–off–нейроны). Нейроны ЯЗТ соединены с
вестибулярными ядрами ствола мозга и нижней оливой. По этому пути зрительные
сигналы о движении достигают центральной вестибулярной системы и (по
оливарно–мозжечковым лазящим волокнам) мозжечка. Обе проекции используются для
глазодвигательной регуляции, особенно для управления горизонтальным ОКН.
Непрямая проекция сетчатки в вестибулярные ядра служит также для восприятия
собственного перемещения в пространстве [6, 17–19, 21, 60].
Ретинотопическая организация зрительного
пути
Принцип топологической организации ЦНС, уже знакомый
нам по соматосенсорной и двигательной системам, реализуется и при нейронном
отображении сетчатки в мозгу. Афферентные и многие центральные отделы
зрительной системы характеризуются «ретинотопической»
организацией. Как различные области планеты наносятся на географическую
карту, так и окружающий нас мир (его изображение на сетчатке) упорядочение
отображается в корковых зонах зрительной системы в виде пространственного
распределения возбужденных нейронов. В отличие от обычной карты с единым
масштабом (скажем, 1:100000, т.е. 1 см изображения соответствует 1 км реального
пространства), ретинотопическая проекция сетчатки на центральной зрительной
системе нелинейна: небольшая центральная
ямка проецируется на гораздо более обширную область зрительной коры, чем такой
же по площади участок с периферии сетчатки. Однако, если рассматривать не
просто эту площадь, а плотность колбочек
или ганглиозных клеток, которая уменьшается от центральной ямки к
периферии, топологическая проекция окажется более или менее линейной. Например,
число ганглиозных клеток сетчатки, связанных с одной корковой колонкой нейронов
(т.е. набором клеток, расположенных перпендикулярно поверхности мозга)
диаметром 1 мм, для всех участков сетчатки примерно одно и то же. Поскольку в
центральной ямке плотность ганглиозных клеток выше, проекция этой области
занимает сравнительно большую площадь корковой зоны VI [17, 18, 54, 56].
Обработка сигналов в латеральном
коленчатом теле (ЛКТ)
Как и ганглиозные клетки сетчатки, большинство
нейронов ЛКТ имеют концентрически организованные рецептивные поля.
Классификация этих клеток примерно такая же, как в сетчатке. Часть их связана с
механизмом усиления одновременного контраста («контрастовые нейроны»), другие
передают преимущественно локальные значения света и тени зрительного стимула («нейроны темного–светлого») или же
отличаются цветоспецифичными рецептивными полями. Парвоцеллюлярные слои ЛКТ состоят главным образом из клеток
последнего типа, а магноцеллюлярные включают
нейроны с рецептивными полями, чувствительными к контрасту и движению. Нейроны
ЛКТ связаны не только с аксонами зрительного нерва; они образуют также
многочисленные синаптические контакты с аксонными окончаниями клеток ствола мозга. Считается, что
именно через эти «незрительные» синапсы на обработку зрительной информации в
ЛКТ влияют степень бодрствования,
пространственно направленное внимание и связанные с ним движения глаз.
Обработка сигналов в первичной
зрительной коре (VI, поле 17)
По нескольким афферентным каналам в первичную
зрительную кору (VI) одновременно
поступает информация о различных свойствах зрительного стимула, например его цвете, контрасте и движении. Аксоны зрительной лучистости образуют синапсы главным
образом на нервных клетках цитоархитектонических слоев IVa–IVc, хотя
некоторые аксонные коллатерали доходят и до нервных клеток других слоев (рис.
11.28). Микроэлектродные записи активности зрительной коры кошек и обезьян
показали, что у нейронов «зернистых» слоев IVa, в, с преимущественно «простые» рецептивные поля. У них либо концентрическая
организация, как в ЛКТ, либо есть параллельно расположенные оп–и off–зоны (рис.
11.29, А). В области VI приматов
обнаружены дополнительные участки, в пределах которых нервные клетки всех слоев
содержат большое количество цитохромоксидазы и имеют простые, в основном
концентрические рецептивные поля. Эти нейроны очень хорошо реагируют на крупные
диффузные световые стимулы и являются преимущественно цветоспецифичными (рис. 11.28: красно–зеленая и сине–желтая
системы). Расположенные снаружи от этих богатых цитохромоксидазой «пузырьков»
нервные клетки слоев I–III и V особенно хорошо реагируют на контуры определенной
ориентации.

Рис. 11.28.
Схема «горизонтальных цитоархитектонических слоев» (I–VI),
глазодоминантных и ориентационных колонок зрительной коры (поле 17=VI) макака–резуса (по [56, 57], схематизировано). В
пределах глазодоминантных колонок ориентационные колонки последовательно
повторяются (изображена только одна такая последовательность). Между колонками
нейронов с рецептивными полями, соответствующими предпочтительной ориентации
контура, находятся более крупные области (также в форме колонок) с нейронами,
рецептивные поля которых лишены такого предпочтения. Эти нейроны реагируют на
диффузные световые стимулы. Одна из их функций–передача цветоспецифичной
информации. Их рецептивные поля организованы концентрически
Таким образом, один из принципов функциональной
организации зрительной коры–расположение
клеток слоями, параллельными ее поверхности.
Как и в других корковых зонах, здесь наблюдается также
второй принцип –геометрическая
организация, перпендикулярная поверхности мозга. Это можно описать
следующим образом.
1. Рецептивные поля нейронов в пределах «корковой
колонки» расположены в одной и той же части сетчатки. Нейроны внутри колонки
связаны друг с другом интернейронами.
Рецептивные поля нейронов соседних колонок перекрываются лишь частично.
2. Нейроны в
колонке диаметром примерно 1 мм возбуждаются в основном сигналами либо от
левого, либо от правого глаза
(«глазодоминантные колонки»). В переходных зонах между соседними
глазодоминантными колонками находятся нейроны, которые примерно одинаково
активируются сигналами от обоих глаз
(бинокулярная интеграция).
3. Глазодоминантные колонки подразделяются на
несколько «ориентационных» (рис. 11.28). У рецептивных полей всех нейронов в
них сходная функциональная «осевая ориентация». Другими словами, лучше всего
они отвечают на контуры одной и той же пространственной ориентации. РП нейронов
в соседних «ориентационных колонках» обычно

Рис.11.29.
Импульсация одиночных нейронов зрительной коры. А. Нейрон с простым
рецептивным полем, у которого on– и off–зоны
расположены параллельно. Б. Нейрон со
сложным рецептивным полем. Его
максимальное возбуждение вызывается смещением в РП наклонной полосы света
ограниченных размеров. В. Нейрон со сверхсложным рецептивным полем. Его
максимальное возбуждение вызывается пересечением под прямым углом двух границ
между темным и светлым. На всех рисунках стимул показан белым. На 5 и 5 стрелки показывают направление его движения (по
данным Hubel, Wiesel,
схематизировано)
различаются по предпочтительной ориентации стимула
примерно на 30°.
4. В других колонках у рецептивных полей нет нейронов
ориентационной специфики; тем не менее, они обычно дают цветоспецифичные ответы
[11, 54–57].
Примеры «отбора сигналов» нейронами
зрительной коры
Рецептивные поля некоторых нейронов в зонах VI и V2 затылочной коры организованы не так, как у
ганглиозных клеток сетчатки и нейронов ЛКТ. У других корковых нейронов они
концентрические и, следовательно, все еще отвечают на диффузные световые
стимулы, однако у множества клеток в коре их on– и off–зоны расположены параллельно (рис. 11.29, А).
Диффузное освещение всего рецептивного поля обычно вызывает лишь слабое
изменение спонтанной активности этих нейронов.
С другой стороны, если на такое рецептивное поле
проецируется полоска света с «правильными» ориентацией и положением,
наблюдается ярко выраженный ответ (рис. 11.29, А). Если полоска света перпендикулярна данному оптимальному направлению, нервная клетка, как правило, не
реагирует. Поскольку параллельные on– и
off–зоны РП сравнительно просто закартировать, проецируя на сетчатку мелкие
световые точки, такие рецептивные поля называют «простыми». Организация on– и
off–зон объясняет, почему эти нейроны дают особенно сильные ответы на контуры определенной ориентации (рис. 11.29, А).
У других нейронов первичной и вторичной зрительной
коры рецептивные поля «сложные».
Мелкие точки света, проецируемые на такие РП, обычно не вызывают выраженной
активности нейронов, в то время как контуры определенных размеров со
специфической ориентацией оказываются наиболее эффективными стимулами, так же
как и разрывы контуров и углы (рис. 11.29,Б, В). Та часть РП, в которой «правильный» стимул вызывает активацию,
называется возбудительным рецептивным
полем (ВРП). Обычно ВРП окружена зоной, в которой светло–темные стимулы
вызывают только торможение (тормозное
рецептивное поле, ТРП).
Корковые нейроны со сложными рецептивными полями
гораздо сильнее активируются не неподвижными, неизмененными в течение
нескольких секунд, а движущимися
стимулами или быстрой сменой
последних. Нейронная активация также частично зависит от направления движения стимула. Чувствительность корковых нейронов к
движению и его направлению позволяет нервной системе решать важнейшую задачу:
«мозговой образ» неподвижного видимого мира должен сформироваться в короткие
периоды фиксаций между саккадами, сдвигающими изображение окружающего
пространства по сетчатке.
Бинокулярная суммация. Обычно,
фиксируя объект окружающего мира двумя глазами, мы видим, что он один. Поэтому
неудивительно, что некоторые нейроны зрительной коры возбуждаются сигналами от
обоих глаз. В основе бинокулярной интеграции лежат различные нейронные
механизмы.
1. Более сильное возбуждение вызывает бинокулярная, а
не монокулярная стимуляция (бинокулярная суммация), причем некоторые из таких нейронов
активируются только при одновременном
возбуждении ВРП сетчаток левого и правого глаза. Если оптимальная ориентация
контура в ВРП левого и правого глаза несколько различна, бинокулярная стимуляция наиболее эффективна, когда контур наклонен
под некоторым углом к фронто–параллельной плоскости и проецируется на
рецептивные поля обоих глаз. Следовательно, эти нейроны участвуют в стереоскопическом восприятии глубины
пространства.
2. Некоторые нейроны в зоне VI возбуждаются при
стимуляции одной сетчатки и тормозятся при стимуляции другой (бинокулярное
торможение). Данный механизм, обнаруженный даже у ряда нейронов ЛКТ (где
бинокулярное торможение передается интернейронами от одного слоя клеток к
другому), составляет нейронную основу «соперничества полей зрения». Считается,
что бинокулярное торможение играет важную роль при тонкой настройке положений
осей глаз, необходимой для бинокулярного зрения.
Зрительные области коры мозга за
пределами первичной зрительной коры (экстрастриарная зрительная кора)
Выполненные сравнительно недавно систематические
микроэлектродные записи активности коры головного мозга обезьян резусов
подтвердили высказывавшуюся ранее гипотезу (основанную на клинических
наблюдениях больных с поражениями мозга), согласно которой зона VI не единственная
корковая зрительная область. У обезьян в обработке зрительной информации
участвуют также обширные зоны затылочной, затылочно–теменной и височной коры
(рис. 11.30). Если у человека стриарная
область (VI) расположена преимущественно на медиальной поверхности
затылочной коры (рис. 11.27, рис. 6.4), то у обезьян она охватывает
значительную часть латеральной затылочной поверхности. Вблизи полулунной борозды она соприкасается в
височном и теменном направлениях с зоной V2 (поле 18). Та в свою очередь ограничена
с тех же сторон зрительными зонами V3, V3a и V4. Эти четыре «экстрастриарные» корковые области
свидетельствуют о специализации нейронной
обработки сигналов в соответствии с разными качественными компонентами зрения.
Нейроны зоны V2 реагируют преимущественно на контуры определенных ориентации и на разрывы контуров. Очевидно, эта зона обеспечивает распознавание
зрительных образов. У нейронов зоны V4
преимущественно цветоспецифичные
рецептивные поля. Поражение этой области у человека приводит к корковому дефициту цветового зрения.
Двустороннее разрушение значительной части «экстрастриарных» зрительных полей
вне зоны VI приводит к зрительной
агнозии. Больные с такой патологией могут «видеть» объекты, но не способны
идентифицировать их по виду, хотя им удается сделать это,

Рис. 11.30. А. Распределение зрительных зон в коре
головного мозга макака–резуса. Область V1 (поле 17) занимает значительную часть наружной
поверхности затылочной доли. Зона с горизонтальной
штриховкой включает части зрительных областей V2, V3, V4, МТ и MST, выходящие на поверхность. Во всех этих
зрительных областях есть упорядоченная проекция сетчатки («ретинотопическое»
отображение), отсутствующая в зрительных областях нижней височной извилины (заштриховано
в клетку). В последней находятся нейроны, отвечающие только на очень
сложные зрительные стимулы. Например, в глубине верхней височной борозды вблизи
вершины соответствующей стрелки есть
клетки, реагирующие преимущественно на лица или лицеподобные фигуры. Б. Горизонтальное поперечное сечение
затылочной коры макака–резуса на уровне, указанном на А стрелками. Различные корковые области обозначены особыми
символами (по [64] с изменениями)
используя другие модальности (например, на ощупь). У
РП нейронов зоны V3 и других зрительных интегративных зон, называемых МТ и MST
(кора вдоль верхней височной борозды), высока чувствительность к движению. Некоторые из этих клеток реагируют
только на движение объектов вдоль оси Z, т.е. к
наблюдателю или от него. В зонах МТ и MST ретинотопическая организация грубая:
у многих нейронов здесь РП очень крупные (до 30°). Двустороннее разрушение этой
области нарушает зрительное восприятие движения [42–45].
Зрительные интегративные области нижней
височной извилины также
отличаются грубой ретинотопической организацией. Здесь находятся нейроны,
реагирующие только на очень сложные зрительные образы. В некоторых случаях их
ответы зависят от процессов визуального научения и текущей мотивации
экспериментальных животных.
Зрительная интегративная область, находящаяся у
обезьяны глубоко в коре вокруг верхней
височной борозды, состоит из нервных клеток, которые легче всего
активируются изображениями лиц или «лицеподобными» стимулами (рис. 11.31).
Некоторые из этих «лицеспецифичных» нейронов дают максимальный ответ на лицо в
профиль, тогда как другие–на «анфас». К зонам, куда направляются их проекции,
относятся части миндалевидного ядра, также
содержащие популяции лицеспецифичных нейронов. Возможно, что. последние,
находясь в области, относящейся к лимбической системе, оценивают информацию об
эмоциональных компонентах выражения лица и движениях его мимических мышц. У
человека «лицеспецифичная» зрительная интегративная область расположена на
базальной границе между затылочной и височной долями. Ее двустороннее поражение
(например, при нарушении кровообращения по ветвям задней мозговой артерии) вызывает симптомы прозопагнозии. Такие больные перестают зрительно различать лица
(Речь идет об узнавании знакомых лиц (близких, родственников и т.д.), а не лиц
«вообще». Последняя способность при прогнозировании сохраняется) или их
выражение, хотя без труда воспринимают другие предметы и могут моментально
узнать знакомых им людей по другим признакам, например голосу или характерным
движениям [40, 50, 59а].
Движение глаз и активация корковых
зрительных нейронов
Как уже упоминалось в начале этой главы, зрительное
восприятие тесно связано с движениями глаз и головы, изменяющими направление
взгляда. Поэтому неудивительно наличие множества различных соединений между
корковыми зрительными областями и глазодвигательными центрами в стволе мозга.
Так, сигналы из области V2 идут в верхние холмики четверохолмия, которые
управляют контролирующими направление взгляда нейронами в ПМРФ и РФСМ.
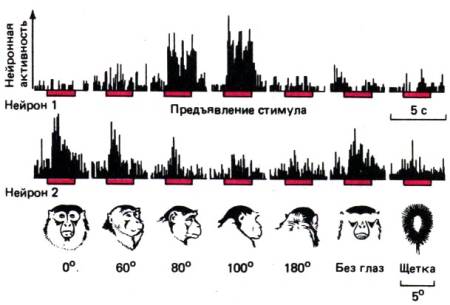
Рис. 11.31.
Средняя частота импульсации (ось ординат)
двух нейронов «лицеспецифичной» корковой области в глубине верхней височной
борозды (см. рис. 11.30). Бодрствующим животным предъявлялись стимулы,
показанные в нижнем ряду (2,5с каждый –красные
горизонтальные полоти). Нейрон 1 дает максимальный ответ на стимул угловым
размером 5°, соответствующий профилю обезьяны, а нейрон 2 на лицо «анфас». Если
закрыть горизонтальную часть лица на уровне глаз, реакция нейрона 2 меняется
незначительно. С другой стороны, предъявление щетки вызывает слабый его ответ
(по [40] с изменениями)
Экстрастриарные зрительные области связаны с
префронтальными корковыми зонами, которые отвечают за произвольные движения
глаз (поле 8) и за координацию движений кисти и рта. В свою очередь, эти
области, подобно полю 7 теменной коры
(см. ниже), соединены с
глазодвигательными центрами ствола мозга и–обратными связями с затылочной
зрительной корой. Путь из сетчатки через нейроны верхних холмиков важен для
афферентной передачи зрительной информации о движении в поле 7. Нейроны верхних
холмиков посылают сигналы в подушку таламуса, откуда они идут к экстрастриарным
корковым зрительным областям и полю 7 теменной коры. Эти связи также
способствуют интеграции движений взгляда и зрительного восприятия.
Нейронное отображение окружающего
пространства
Координаты и пространственная организация объектов в
окружающем мире кажутся нам неизменными, даже когда их изображения на сетчатке
смещаются при активных движениях глаз, головы и тела. Более того, по крайней
мере в той части внешнего пространства, которая находится в пределах
непосредственной досягаемости, предметы имеют мультимодальные свойства: чашка
воспринимается как таковая, когда мы смотрим на нее, когда прикасаемся к ней и
когда из нее пьем. Обширные поражения теменной коры головного

Рис. 11.32.
Схема ответа нейрона поля 7 бодрствующего макака–резуса. А. Объект (например, орех) находится в пределах досягаемости и
движется; движение возбуждает нейрон (частота его импульсации увеличивается). Б. Животное смотрит на объект; частота
импульсации возрастает еще больше, В.
Когда животное хватает объект, возбуждение нейрона достигает максимума [47]
мозга человека нарушают восприятие предметов в
пространстве или полностью препятствуют ему. Как правило, этот дефицит
ограничен контралатеральной месту поражения половиной пространства
(одностороннее зрительное
игнорирование). (У правшей встречается преимущественно синдром
«игнорирования левой половины пространства» при поражениях теменной доли
правого полушария. Многие авторы специально выделяют его как показатель
поражения именно правого полушария). Обычно он сопровождается нарушением
произвольных движений глаз в контралатеральную сторону. Функцию теменных полей
коры изучали с помощью микроэлектродов, вживленных в мозг обезьяны. В поле 7 здесь
находятся нейроны, возбуждаемые как зрительными, так и тактильными стимулами.
Их активность частично зависит от
движений глаз, хотя на нее влияют также интерес и та часть окружающего
пространства, на которое животное обращает внимание (рис. 11.32). Очевидно, что
в поле 7 отображение окружающего не зависит от ретинотопической проекции.
Зрительные области подушки, посылающие сигналы к полю 7, также участвуют в
обеспечении этих сложных функций [5, 39, 48, 58, 59].
11.6.
Практические и клинические аспекты физиологии зрения
Нейрофизиологические основы восприятия
формы
Хороший рисовальщик способен несколькими штрихами
передать «суть» сложного объекта, отбросив несущественные детали и выделив главное
в структуре изображаемого. Зрительные нейронные сети в коре головного мозга
занимаются такого же рода «выделением». Нейронные операции, направленные на
выявление отдельных признаков–контуров с определенной ориентацией или длиной,
разрывов линий или углов контура,–можно рассматривать как первый этап этого
процесса. В каждую сеть входит специфический класс нейронов со сложными
рецептивными полями; характерному пространственному распределению их
возбуждения соответствует тот или иной признак формы стимула. В упрощенном
примере на рис. 11.33 это иллюстрируется для стимула в виде буквы «К». Однако пока неизвестно, какие нейронные механизмы
интегрируют многочисленные нейронные отображения в зрительной коре, обеспечивая
восприятие целостного объекта (в данном случае буквы «К»).
Зрительные иллюзии.
Некоторые хорошо известные зрительные иллюзии можно объяснить организацией
рецептивных полей корковых нейронов. Например, непредвзято рассматривая рис.
11.34, А, мы увидим белый квадрат на
темном фоне. Как показывают микроэлектродные записи, в области V2 есть нейроны со сложными рецептивными полями,
реагирующие на «кажущиеся контуры». В таком случае весьма вероятно, что
восприятие кажущихся контуров на рис. 11.34, А вытекает из организации
рецептивных полей этих клеток.
Иллюзия Мюллера – Лайера (рис. 11.34,Б),
видимо, также объясняется организацией сложных рецептивных полей корковых
нейронов. Как уже говорилось выше, возбуждение некоторых из них определяется
углом между двумя контурами, а этот параметр весьма важен для эффективности
данной иллюзии. Зрительные иллюзии–особые случаи восприятия формы, помогающие
физиологу, изучающему сенсорные системы, понять ход обработки информации в
зрительных центрах мозга.
Фигура и фон при восприятии формы. Различные фигуры можно видеть только тогда, когда они
выступают из «фона». В определенных условиях разница между фигурой и фоном
бывает неоднозначной. Это иллюстрирует рис. 11.34,5. На нем можно различить
либо силуэт черного кубка (или подсвечника) на белом фоне, либо два белых улыбающихся
друг другу лица в профиль на черном фоне. Одновременно
воспринимать обе эти формы невозможно. Если долго смотреть на изображение,
фигуры и фон неизбежно будут чередоваться, видимо, из–за процесса адаптации
зрительной системы за пределами зоны VI.

Рис. 11.33.
Схема распределения возбуждения, вызываемого светящейся буквой «К» (см. рис.
11.5) в разных нейронных, слоях сетчатки и в центральной нервной системе, а. Изображение буквы на сетчатке и
пространственное распределение возбуждения в ее рецепторном слое. б. в. Возбуждение на выходе из сетчатки
(в слое ганглиозных клеток) (на рис. б–и
возбужденные области показаны красным), б.
Нейроны с оп–центрами. в. Нейроны с
off–центрами. г. Распределение
возбуждения в нейронах латерального коленчатого тела и слоя IV зрительной коры
(с концентрическими РП). Нейроны активируются контурами буквы, д–и. Возбуждение в разных нейронных
слоях и типах клеток первичной, вторичной и третичной зрительной коры. Эти
нейроны активируются только контурами определенной ориентации, с определенными
углами или разрывами. Данная схема сильно упрощает реальную картину. На самом
деле пространственное распределение возбуждения в корковых нейронных слоях
отличается от линейного преобразования стимула
То же самое справедливо и в отношении пространственной
неоднозначности «куба Неккера» на рис– 11.34, Г. Если смотреть на него
некоторое время (одним глазом или обоими), его ориентация покажется изменчивой
–ребро «а» оказывается расположенным то спереди, то сзади. Можно произвольно
заставить куб Неккера не «перепрыгивать» из одного положения в другое, однако
«удержать его на месте» удается не более чем на несколько секунд.
Постоянство
формы и размера. Как правило, мы воспринимаем окружающие предметы
неизменными по форме и размеру, хотя на самом деле их угловые размеры и форма
изображений на сетчатке непостоянны. Например, велосипедист в поле нашего
зрения всегда кажется одинаковым по величине, независимо от расстояния до него.
Колеса велосипеда воспринимаются как круглые, хотя на самом деле их изображения
на сетчатке могут стать узкими эллипсами. Это явление постоянства формы и
размера демонстрирует роль опыта в видении окружающего. Он влияет и на
восприятие глубины внешнего пространства. Область в пределах нашей
непосредственной досягаемости и поблизости от нее, доступная для взаимодействия
с окружающими предметами, является
покажутся игрушечными, однако, если рассматривать
такие же объекты на расстоянии 150 м по горизонтали, они сохранят свой
«истинный» размер. Нелинейное искажение дальнего видимого пространства причина
так называемой «лунной» иллюзии: луна или солнце кажутся гораздо мельче в
зените, чем у горизонта. Говорят, что, если долго работать на высоте, такое
неевклидово искажение пространства изменяется. При повреждении теменнозатылочной
коры головного мозга может пострадать механизм, контролирующий постоянство
воспринимаемого размера. В этом случае предметы могут казаться аномально
мелкими (микропсия), слишком
крупными (макропсия) или искаженными (дисморфопсия) [9, 12, 13, 30].
Измерение остроты зрения
При обычном положении глаз изображение фиксируемого
предмета проецируется в их центральные ямки. В условиях фотопического зрения
это место сетчатки соответствует максимальной остроте зрения; к периферии
сетчатки она снижается (рис. 11.35). При скотопическом зрении острота зрения
максимальна в области, окружающей центральную ямку. Там, где из глаза выходит
зрительный нерв, острота зрения нулевая (слепое пятно, рис. 11.4).
При клиническом определении «визуса» (V), т. е.
остроты зрения в точке, где она максимальна, пациента обычно просят фиксировать
стандартную цель. Наиболее распространенный способ измерения остроты зрения в
США и некоторых странах Европы–вычисление отношения Снеллена
V=d/D, (11)

Рис. 11.34. А. Пример зрительного «достраивания»
формы (белый квадрат). Б. Иллюзия
Мюллера–Лайера: на самом деле длина двух вертикальных линий одинакова. В. Рисунок, демонстрирующий чередование
фигуры и фона. Наблюдатель видит либо черный «подсвечник» на белом фоне, либо
белые профили двух улыбающихся людей на черном фоне. Г. Куб Неккера. Если смотреть достаточно долго, его
пространственные соотношения меняются–задние ребра «перепрыгивают» вперед, а
передние–назад. Такие перескоки восприятия нельзя подавить сознательно
«евклидовой», т. е. равные по длине отрезки в ней
воспринимаются одинаковыми по всем направлениям. Однако для более дальнего
окружения это не так. Большинство читателей может убедиться в этом на
собственном опыте. Если смотреть с высоты 100–150 м вниз, люди и машины там где
d– расстояние, на котором данный стимул различается, а
D–расстояние, соответствующее его угловому размеру 1’. В данном случае
символами служат буквы таблицы Снеллена. Ее помещают на стандартном расстоянии
от пациента (обычно 6,5 м), а D вычисляют по размеру наименьших букв, которые
он может прочитать. Для людей с нормальным зрением D и d одинаковы и отношение
Снеллена равно единице.
Еще один способ–определение угловой величина α
различимого пациентом разрыва в кольце Ландольта (рис. 11.35). В данном случае
острота зрения вычисляется по формуле V = 1/ α и измеряется в (угл. мин)–1.
Тогда нормальный визус (различение цели размером 1’) составляет 1/угл. мин.
Если остроту зрения определяют после коррекции с помощью очков аномалий
лучепреломления, полученное значение визуса называют visus con correctione (visus с. с.);
визус, измеренный без очков,–visus
sine correctione
(visus s. с.)
[20, 25, 26].
Определение поля зрения и его дефектов с
помощью периметрии
Монокулярное поле зрения представляет собой часть пространства, воспринимаемую
неподвижным глазом. Общее поле зрения
включает все точки, видимые двумя
Рис. 11.35.
Зависимость остроты зрения (ось ординат)
от положения стимула в поле зрения (ось абсцисс).
Черная кривая –фотопическое зрение, красная
скотопическое. Острота зрения измерялась с помощью кольца Ландольта (на
врезке). Данный рисунок можно использовать для демонстрации наличия слепого
пятна. Для этого нужно правым глазом фиксировать крест (буква F) с расстояния примерно 32 см. В этом случае кольцо
Ландольта проецируется на область слепого пятна и становится невидимым
неподвижными глазами. В пределах общего поля зрения выделяется область, которую видят два
глаза одновременно,–бинокулярное поле зрения. С каждой его стороны располагается
область, видимая одним глазом,–монокулярный полумесяц. Разумеется, за счет
движений глаз даже при неподвижной голове человек воспринимает область,
превышающую общее поле зрения (поле взгляда).
Дефект поля зрения–это утрата зрительных ощущений в
какой–либо его части. Если дефектная зона окружена со всех сторон нормальным
полем зрения, она называется скотомой.
Такие дефекты бывают связаны с поражениями либо сетчатки, либо центральной
зрительной системы. Границы нормального или аномального поля зрения и положение
скотом картируют с помощью периметрии (рис.
11.36, 11.37). Для проведения точной периметрии должны быть строго определены
такие параметры, как характер адаптации глаз испытуемого, размер, интенсивность
и спектральный состав светового стимула. По характеру дефектов поля зрения врач
может установить место поражения зрительных путей (если, конечно, он хорошо
знаком с анатомическими взаимоотношениями, рассмотренными выше. При обычных
измерениях (взгляд направлен прямо вперед) поле зрения с одной стороны
ограничено носом. Чтобы обследовать носовую область поля зрения, испытуемого
просят отвести взгляд вбок от стандартного положения. У светоадаптированного
человека поле зрения для черно–белых стимулов шире, чем для цветных (рис.
11.36). Функциональная «цветовая слепота» его периферии объясняется слишком
малым числом колбочек в этой же части сетчатки [3, 24, 30].
Электрофизиологическое изучение
зрительной системы
Работу зрительной системы можно проверить объективными
методами–на сетчаточном или центральном уровнях–с помощью
электрофизиологических измерений. Выше уже упоминалась регистрация электроретинограммы. Другой важный
объективный способ проверки афферентной части зрительной системы–запись зрительных вызванных потенциалов
(ЗрВП) при электроэнцефалографии (ЭЭГ) с помощью электродов, помещенных на кожу
головы над мозговой коробкой. ЗрВП получают, суммируя реакции на неоднократно
предъявляемые стимулы. Вспышка рассеянного света в обширной области дает
простой ЗрВП; его волны и латентные периоды зависят от яркости и спектрального
состава стимула (рис. 11.38, А). ЗрВП обращаемого узора измеряют с
помощью рисунков в виде черных и белых
полос или шахматных клеток, светлые
и темные участки которых периодически меняются местами (рис. 11.38, .6). Если
же стимулы различной конфигурации сменяют друг друга апериодически, ЗрВП дает
объективную информацию о более сложных процессах обработки информации в ЦНС
(рис. 11.38, В, Г). При множественном
склерозе–одном из наиболее часто встречающихся неврологических заболеваний–изменяются
латентные периоды и амплитуды волн ЗрВП обращаемого

Рис. 11.36. А. Схема аппарата для периметрии. Поле
зрения (5) оценивается монокулярно. Испытуемый располагается перед аппаратом
таким образом, что его глаз совпадает с центром полусферы и фиксирует взглядом
точку на ее полюсе (О). Врач проверяет правильность фиксации через специальное
отверстие и перемещает световое пятно по поверхности сферы прибора с помощью
проекционной системы, управляемой ручкой Р. Световое пятно может иметь разную
величину, яркость и цвет. Как только испытуемый замечает пятно, он подает
сигнал, и это положение пятна регистрируется на бумаге, закрепленной на
подставке П. Б. Результат определения
границ нормального поля зрения для белого, синего и красного стимулов.
СП–слепое пятно. Точка фиксации соответствует центру концентрических кругов,
обозначающих удаленность стимула от точки фиксации (в угловых градусах)
узора, поскольку при этой патологии обычно
повреждаются аксоны зрительного нерва (ретробульбарный неврит). Таким образом,
измерения ЗрВП–важный диагностический метод в клинической неврологии и
нейроофтальмологии [28].

Рис. 11.37.
Схема типичных дефектов поля зрения. Возможные пораженные участки: a –сетчатка правого глаза (область центральной ямки)
или правый зрительный нерв; б–зрительный перекрест; в левый зрительный тракт
или центральные отделы левого полушария мозга; г–сетчатка левого глаза или
левый зрительный нерв; д– правый зрительный тракт или
центральные отделы правого полушария мозга. Локализация этих поражений показана
на рис. 11.27
Восприятие глубины
В местности с хорошо выраженным рельефом человек с
нормальным бинокулярным зрением воспринимает детали ландшафта как расположенные
на разных расстояниях от него. При этом он может довольно точно определить
относительную удаленность тех или иных объектов. Такое ощущение глубины
пространства обусловлено отчасти бинокулярным стереоскопическим зрением. Однако
определенную роль здесь играют и монокулярные
механизмы: различия в размерах знакомых
предметов, перекрывшие деталей, отбрасываемые тени, перспективное укорочение
и особенно параллакс, т. е. смещение одних предметов относительно других
при движениях головы наблюдателя. Трехмерное восприятие объектов с помощью бинокулярной стереоскопии особенно
важно, когда они расположены в зоне непосредственной досягаемости или вблизи
нее. Поскольку глаза расположены в разных местах головы, изображения ближних
объектов на их сетчатках несколько различны. Это следует из законов
геометрической оптики. Чем крупнее предметы и чем ближе они к наблюдателю, тем
больше такое горизонтальное расхождение
(диспаратность).
То, что изображения одного объекта на двух сетчатках
различны, легко показать с помощью простого эксперимента. Вытяните руку перед
собой и рассматривайте

Рис. 11.38.
Зрительные вызванные потенциалы, записанные у человека от
затылочной области поверхности головы. Каждая кривая усредненный ответ на 40
стимулов. А. Реакция на диффузную
вспышку света длительностью 20 мс, освещающую участок бинокулярного поля зрения
размером 5 х 7°. Простой вызванный потенциал. Б. Потенциал обращаемого узора. Испытуемый видит черно–белый узор
из вертикальных полос, в котором в момент, указанный стрелкой, белые полосы меняются местами с черными. Средняя яркость
стимула не меняется. Его размер 10х15°, период повторения полос 2°. В. Потенциал, вызываемый изменением
формы (этот момент указан стрелкой).
Изображение лица предъявляется после рисунка дерева или стула. Г. В момент, указанный стрелкой, изображение дерева
предъявляется после рисунков лица или стула. «Формозависимые» вызванные
потенциалы (В, Г) различаются ходом
волн. Наибольшее различие по напряжению между двумя кривыми наблюдается в
момент, соответствующий 220 мс (измерения К. Botzel, Берлин)
большой палец монокулярно то правым, то левым глазом.
При каждой смене глаза создается впечатление, что палец скачком перемещается
относительно фона. Другими словами, когда на палец смотрит правый глаз, тот
закрывает одни предметы, когда левый–другие. Чем дальше объекты заднего плана
от руки, тем больше смещение относительно них пальца. Геометрическая схема
возникновения горизонтальной диспаратности показана на рис. 11.39. Когда оба
глаза фиксируют точку на определенном расстоянии, все объекты на окружности,
проходящей через узловые точки оптических систем двух глаз и точку фиксации,
проецируются на корреспондирующие
участки сетчатки. Все объекты внутри или за пределами этой окружности (гороптерного круга) проецируются на
некорреспондирующие участки. Если горизонтальная диспаратность (сумма углов
α и β на рис. 11.39) достаточно велика, изображение двоится. Пространственное разделение
двух изображений при горизонтальной диспаратности можно измерить, использовав
модель «циклопического глаза».
Горизонтальная диспаратность и
бинокулярное слияние.
Итак, когда голова наблюдателя находится в обычном
положении, изображения объекта, расположенного на конечном расстоянии от глаз,
проецируются в разные места их сетчаток. Количественная мера этой
разницы–горизонтальная диспаратность. Когда она превышает определенную
величину, бинокулярно наблюдаемый трехмерный предмет воспринимается двоящимся.
Диапазон расстояний до объекта–от нулевой диспаратности (объект в
бесконечности) до предела бинокулярного глубинного зрения, при котором
наступает двоение, весьма велик. Это можно объяснить, исходя из величины
рецептивных полей бинокулярно активируемых корковых нейронов: размер ВРП таких
нейронов в областях VI и V2 значительно
больше, чем пространственное разрешение соответствующего участка сетчатки. У
некоторых из корковых нейронов РП на обеих сетчатках точно совпадают. По этой
причине они сильнее всего возбуждаются объектами, расположенными в пределах
гороптера. С другой стороны, РП некоторых бинокулярно активируемых нейронов
точно на сетчатках не совпадают. Эти клетки сильнее всего возбуждаются
объектом, находящимся на определенном расстоянии от гороптера, что приводит к
некоторой горизонтальной диспаратности.
Стереоскопическое восприятие глубины важно в некоторых
областях медицины, особенно в хирургии, главным образом при микрохирургических
операциях с использованием бинокулярного микроскопа. Врачам, не обладающим
нормальным бинокулярным зрением, трудно проводить их, поэтому они должны
избегать подобной специализации. Стереоскопическое зрение легко проверить,
попросив испытуемого посмотреть бинокулярно на два находящихся рядом рисунка,
выглядящие для каждого глаза по отдельности как случайные наборы точек. Только
те, кто способен бинокулярно коррелировать отдельные точки с соответствующей
горизонтальной диспаратностью, воспринимают в данном случае форму и глубину
изображений [12, 13, 14, 16, 25].
Зрительное восприятие движения и
собственного движения наблюдателя
Восприятие глубины пространства и расстояния за
пределами зоны непосредственной досягаемости основано главным образом на
параллаксе изображений на сетчатке при перемещении наблюдателя. Если равномерно
двигаться вперед, например в автомобиле или при взлете и снижении самолета,
смещение изображений на сетчатке будет описываться простым правилом: чем ближе объект в поле зрения, тем больше
его угловая скорость. Такое кажущееся движение можно имитировать с помощью
последовательности изображений, заснятых на пленку или синтезируемых на экране
компьютера. Если неподвижно сидящий человек смотрит такой

Рис. 11.39.
Схема бинокулярного зрения и построения «циклопического глаза». Когда объект
расположен снаружи от гороптера, его изображение проецируется на участок справа
от центральной ямки в левом глазу и слева от нее в правом. В этом случае при
бинокулярном зрении формируются его непересекающиеся двойные изображения. Их
положение можно определить, проецируя сетчатку правого и левого глаза на
воображаемую сетчатку циклопического глаза
широкоформатный фильм, ему кажется, что он движется
навстречу потоку оптических сигналов
(перенос). Кажущееся собственное
движение может иметь линейную или вращательную составляющие (линейный и круговой переносы
соответственно). К примерам такого восприятия относятся ощущения, переживаемые
на авиационном или автомобильном тренажере, а также при езде на транспорте с
постоянной скоростью. Чем реалистичнее воссоздает паралакс последовательность
изображений, т.е. чем лучше она имитирует трехмерность зрительно
воспринимаемого пространства, тем естественнее воспринимается кажущееся
движение. Перенос объясняется наличием связей между сетчаткой и вестибулярными
ядрами, а также взаимодействием центральной зрительной системы с вестибулярными
зонами коры головного мозга.
Косоглазие
Если при фиксации объект проецируется на центральную
ямку только одного глаза, а в другом его изображение строится в окружающей ее
области, возникает косоглазие (страбизм).
Если оптические оси глаз достаточно сильно при этом расходятся, изображение
двоится. Это происходит, например, при параличе мышцы одного глаза, нарушающем
координацию его движений со вторым. У таких больных двоение возникает всякий
раз, когда требуется выполнить движение глаз, требующее участия пораженной
мышцы. Чтобы точно определить, какая мышца парализована, необходимо тщательно
описать взаиморасположение двух видимых пациентом изображений (пересекаются или
не пересекаются и т.д., рис. 11.39). Такое
двоение легко воспроизвести, зафиксировав бинокулярно ближний предмет, а затем
немного сместив один глаз пальцем.
Если в раннем детстве бинокулярные движения глаз не
скоординированы (врожденное косоглазие), у одного глаза обычно развивается амблиопия: детальная структура
зрительных стимулов различима только центральной ямкой одного глаза (не амблиопического).
Причина такого нарушения связана не с нарушением работы сечатки, а с
функциональными изменениями зрительной коры; обработка информации, поступающей
от амблиопического глаза, на уровне корковых нейронов ухудшается или полностью
прекращается. Амблиопия развивается также у детей, долгое время вынужденных
смотреть только одним глазом (например, при птозе, т. е. врожденном опускании
верхнего века).
Наиболее частая причина амблиопии при косоглазии у детей–гиперметропия. Самые юные
пациенты, чтобы сделать изображение на сетчатке более резким, при аккомодации
вынуждены сильно напрягаться. Однако при рассматривании близких объектов между аккомодацией и конвергенцией существует
тесная связь, поэтому глаза дальнозорких детей конвергируют на больший угол,
чем требуется на данном расстоянии, и объект оказывается на оптической оси
только одного из них. Упоминавшийся выше бинокулярный тормозный механизм
подавляет сигналы, идущие в зрительную кору от другого глаза. Если вовремя не
скорректировать гиперметропию очками и не вылечить косоглазие, в конечном счете
неизбежно возникнет неизлечимая
амблиопия, поскольку синаптические контакты между корковыми бинокулярными
нейронами и афферентами из ЛКТ, связанными с пораженным глазом, станут
нефункциональными.
Косоглазие вызываются также нарушением глазодвигательной координации в стволе мозга.
Результатом опять–таки будет амблиопия. Лечение косоглазия у детей–с помощью
очков, специальных упражнений для глаз или хирургическим путем–должно быть
проведено до двух–, трехлетнего возраста, поскольку затем «сенситивная фаза»
формирования корковых нейронных механизмов бинокулярного зрения завершается и
лечение амблиопии обычно становится неэффективным. Последующая корректирующая
операция дает лишь косметический эффект.
11.7. Цветовое зрение
Цвет и измерение цвета
Феномен цветового зрения особенно ясно
свидетельствует, что восприятие зависит не только от вида стимулов и работы
рецепторов, но и от характера обработки информации в нервной системе. В
психофизике цветового восприятия выделяют две главные характеристики окраски–ее хроматические (красная, оранжевая,
синяя и т. д.) и ахроматические (от
глубокой черной через различные оттенки серой до самой яркой белой)
«валентности». В видимом солнечном
спектре (например, в случае радуги) различные зоны кажутся окрашенными
по–разному, давая плавный переход цветовых ощущений–от фиолетового через синий,
зеленый, желтый, оранжевый до красного. Однако это не означает, что ощущение
цвета определяется только длиной волны \
монохроматического светового стимула. С одной стороны, мы способны воспринимать
цветовые оттенки, отсутствующие в солнечном спектре, например пурпурный (смешение красного и
синего). С другой стороны, все цвета видимого спектра можно получить путем
смешения света с другими длинами волн. Цвета,
получаемые путем спектрального смешения, неотличимы от чистых спектральных
цветов.
Феноменологическая структура цветовосприятия описывается законами цветового зрения. «Цветовое пространство» нормального
человека содержит примерно 7 млн. различных валентностей, включая небольшую
категорию ахроматических и весьма обширный класс хроматических. Хроматические
валентности поверхностной окраски объекта характеризуются тремя
феноменологическими качествами:
тоном, насыщенностью и светлотой. В случае светящихся цветовых стимулов (например,
цветного источника света) «светлота» заменяется на «яркость».
Цветовые тона
образуют естественный бесконечный континуум, цветовой круг с последовательностью: красный, желтый, зеленый,
синий, пурпурный и снова красный (рис. 11.40, А). В идеале тона–это «чистые»
цвета. Тон может быть «смешан» с ахроматической валентностью, что дает
различные оттенки цвета. Например, чистый красный при смешении с белым дает
розовый, а при смешении с черным – коричневый. Насыщенность оттенка–это мера относительного содержания в нем
хроматических и ахроматических компонентов, а светлота определяется положением ахроматического компонента на
шкале серого.
Все воспринимаемые цветовые валентности можно
представить в виде трехмерного цветового
тела, в котором относительное положение цветов определимо с метрической
точностью или же задается чисто качественно. На рис. 11.40, А 'показан
один из первых (1810 г.), неметрических, вариантов такой формы
представления–цветовой шар немецкого
художника Филиппа Отто Рунге. Каждой цветовой валентности здесь соответствует
определенный участок на поверхности шара или внутри него. Сечение по экватору
дает на периферии чистые цвета цветового круга, которые к центру все больше
смешиваются с серым. Центр шара занимает нейтральный серый цвет. Вдоль оси,
идущей от одного полюса до другого, располагаются все оттенки серого–от белого
до глубокого черного. В современных моделях, где восприятие цвета задано
метрически, простой шар Рунге деформируется в нешаровидное цветовое тело. Цель
создания таких метрических цветовых систем – однозначное описание цветовосприятия здоровых людей. Они не дают
физиологического объяснения
цветового зрения. Однако любая физиологическая теория цветового зрения должна
объяснять структуру метрического психофизического цветового пространства.
Постоянство цвета.
В повседневной жизни важно, чтобы при естественном освещении восприятие цветов
поверхностей было относительно независимым от спектрального состава падающего света.
Чтобы обеспечить такую независимость, объекты должны освещаться светом,
включающим значительный диапазон видимого спектра. Постоянство их цвета связано
с восприятием нами преимущественно
спектральной отражательной способности поверхностей. Она позволяет
распознать объекты в естественных условиях при различном освещении. Однако у
постоянства цвета есть свои границы. Сравните, например, контраст между
коричневым стволом пихты и глубоким зеленым

Рис. 11.40. А. Распределение тонов на цветовом круге.
Тоны между А и Б (стрелка) не
относятся к спектральным, а получаются за счет смешения красного и синего. Б. Цветовой шар Рунге (1810
г.)–неметрическое представление цветового пространства
цветом ее иголок в серый дождливый день с гораздо
более сильным цветовым контрастом в случае, когда дерево освещается в летний
вечер красноватым светом заходящего солнца. При искусственном освещении,
соответствующем только части видимого спектра, цвета тканей кажутся иными, чем
при естественном. Это следует помнить, когда покупаешь одежду.
Смешение цветов.
Из повседневного опыта мы знаем, что при
смешении красок разного тона получается цвет другого тона. Однако
последствия смешения красок совершенно отличны от того, что получается при
сочетании света из разных частей спектра.
При аддитивном смешении цветов свет с
разной длиной волны падает на одну и ту же точку сетчатки (рис. 11.41).
Если отдельные источники света монохроматические, аддитивное смешение может
дать тон, соответствующий другой части спектра или же неспектральной области
между красным и синим (пурпурный). Описание цветовых соотношений в форме
цветового тела частично основано на изучении уравнений аддитивного смешения
цветов. Эти уравнения используются также в тестах по выявлению дефицита цветового зрения. Инструмент
для проведения таких испытаний называется
аномалоскопом. С помощью него можно, например, спроецировать спектральный
желтый

Рис. 11.41.
Схема аддитивного (А) и
субтрактивного (Б) смешения цветов
(λ= 589 нм) на одну половину круга, а смесь спектральных
красного (λ = 671 нм) и зеленого (λ = 546 нм)–на другую (рис. 11.41).
Задача испытуемого – подобрать соотношение красного и зеленого, чтобы
получилась смесь желтого цвета, неотличимая от его спектрального эквивалента.
Результат описывается уравнением смешения цветов
а(красный,
671)+ b(зеленый, 546) ≈(желтый, 589), (12)
где символ ≈ означает эквивалентность ощущения и не имеет математического смысла. У людей
с нормальным цветовым зрением коэффициент для красной составляющей–примерно 40,
а для зеленой–около 33 относительных единиц (если для желтого цвета он равен
100).
Если смешать аддитивно два монохроматических световых
стимула, один в диапазоне 430–555 нм, а другой–492–660 нм, всегда можно
получить белый цвет, подобрав соответствующую их пропорцию. Однако смешение
монохроматических лучей с длиной волны более 660 и менее 430 нм белого цвета
никогда не даст; получатся оттенки пурпурного, отсутствующие в спектре (рис.
11.40, 11.43).
Белый цвет. Для каждого тона на цветовом круге существует другой
тон, дающий при аддитивном смешении с ним белый цвет. Константы (весовые
коэффициенты a и b) такого уравнения смешения цветов
a (F1) + b (F2 ) ≈ с (белый) (13)
зависят от определения белого цвета. Любую пару тонов F1 и F2,
удовлетворяющую уравнению (13), называют
дополнительными цветами.
Субтрактивное смешение цветов отличается от аддитивного тем, что является чисто
физическим процессом.
Если белый свет пропустить через два фильтра с широкой
полосой пропускания–сначала через синий, а затем через желтый,–получится
«смесь» зеленого цвета, поскольку
через оба фильтра могут пройти
только зеленые световые лучи (рис. 11.41). Художник, смешивая краски,
производит субтрактивное смешение цветов, поскольку отдельные гранулы пигментов
действуют как широкополосные хроматические фильтры.
Трихроматичность. При нормальном цветовом зрении любой тон (F4) может
быть получен путем аддитивного смешения трех других определенным образом
подобранных тонов (F1 — F3). Это записывается в виде уравнения необходимых и достаточных
условий цветоощущения
a (F1) + b (F2 ) + с (F3 ) ≈ d (F4 )
(14)
Согласно международной конвенции, в качестве главных
цветов F1, F2 и F3 используемых
для построения современных метрических цветовых систем, выбраны спектральные
цвета с длинами волн 700 (красный), 546 (зеленый) и 435 (синий) нм. Для
получения белого цвета при их аддитивном смешении весовые коэффициенты должны
быть связаны соотношением
a+b+c=d=l. (15)
Результаты физиологических экспериментов по
цветовосприятию, описываемые уравнениями (11)–(14), могут быть геометрически
представлены в виде диаграммы цветности
(«цветового треугольника»), приведенной на рис. 11.42. Оттенки серого,
присутствующие в цветовом теле,
здесь опущены, так что трехмерная модель вырождается в двумерную. Согласно этой
диаграмме, при смешении двух цветов точка, соответствующая получаемому цвету,
лежит на соединяющей их прямой. Чтобы найти пары дополнительных цветов, необходимо провести прямую через «белую
точку» (Е) [9, 12, 13, 16, 31, 33].
Теории цветового зрения. Ниже описаны две наиболее известные из этих теорий. В
свое время между их сторонниками велись жаркие споры, однако сейчас обе можно
рассматривать как взаимодополняющие теоретические интерпретации феномена
цветового зрения. Каждая из них «правильно» описывает свой уровень афферентной
зрительной системы и подтверждается прямыми физиологическими измерениями.
Попытка синтеза двух этих конкурирующих теорий была предпринята 80 лет назад
Иоганнесом фон Крисом, предложившим свою
зонную теорию.
Трехкомпонентная теория, разработанная Юнгом, Максвеллом и Гельмгольцем,
постулирует, что три разных типа колбочек при фотопическом зрении работают как независимые рецепторные системы.
Комбинации получаемых от них сигналов анализируются двумя нейронными системами
– восприятия яркости и цвета.
Справедливость этой теории подтверждается закономерностями цветовосприятия на
нижнем пределе фотопической чувствительности: в таких условиях различаются
только три тона – красный, зеленый и синий. Прямые измерения спектров поглощения
фотопигментов одиночных колбочек и записи их рецепторных потенциалов в сетчатке
животных с цветовым зрением (см. и далее) объективно подтвердили наличие трех
типов цветовых рецепторов.

Рис. 11.42. Цветовой
треугольник, соответствующий немецкому стандарту DIN 5033. Область
белого цвета окружает точку Е. В основании треугольника–оттенки пурпурного
тона. При аддитивном смешении двух цветов А и В результирующий цвет лежит на
линии, соединяющей соответствующие точки. Дополнительные цвета лежат на линиях,
проходящих через точку Е
Теория оппонентных цветов. Полярное расположение тонов на цветовом круге легло в
основу другого подхода к цветовосприятию. Мах и Геринг предположили, что все
хроматические валентности обусловлены четырьмя первичными цветами (Urfarben) –
красным, желтым, зеленым и синим. Согласно Герингу, они связаны попарно двумя
физиологическими процессами, у каждого из которых антагонистическая
организация,–восприятия зелено–красного и желто–синего. Третий оппонентный
процесс постулирован для ахроматических дополнительных цветов–белого и черного.
Термин «оппонентные цвета» основан на полярной природе цветовосприятия. Другими
словами, не может быть «зеленовато–красного» и «синеватожелтого» цветов. Эта
теория подтверждается эффектами одновременного и последовательного
хроматического контраста. Например, если серый круг окружен ярко–зеленым
кольцом, то за счет одновременного цветового контраста он приобретает
красноватый оттенок. Если кольцо убрать, наблюдатель увидит красное кольцо,
окружающее зеленоватый круг (последовательный хроматический контраст). Теория
оппонентных цветов Геринга постулирует наличие антагонистических
цветоспецифичных нейронных механизмов. Вероятно, уровень, на котором они действуют,
у разных позвоночных неодинаков [5, 8, 9, 12, 13, 16, 31].
Физиологическая основа цветового зрения
Микроспектрофотометрия пигментов колбочек и
микроэлектродные измерения мембранного потенциала отдельных колбочек у животных
с цветовым зрением подтверждают предсказания трехкомпонентной теории: имеются
три основные группы рецепторов, различающиеся своими спектрами поглощения (см.
рис. 11.15). Антагонистические реакции, постулируемые теорией оппонентных
цветов, также обнаружены в сетчатке. Речь идет о спектральных ответах
горизонтальных и биполярных клеток, обусловленных пространственным
взаимодействием сигналов от колбочек разных типов. Кривые, приведенные на рис.
11.43,А–В, получены при измерении реакции горизонтальных клеток сетчатки рыбы,
которая, как показали поведенческие эксперименты, обладает хорошим цветовым
зрением. Обнаружено три класса горизонтальных клеток. Один из них под действием
света гиперполяризуется независимо от длины волны (L–mun). Два
других демонстрируют красно–зеленый и
сине–желтый антагонизм (рис. 11.43,Б,
В). Они гиперполяризуются, когда глаз
стимулируется монохроматическим светом одной части спектра, и деполяризуются
под действием света из другой его части. У млекопитающих с цветовым зрением
обнаружена аналогичная цвето–специфичная организация некоторых биполяров с

Рис.
11.43. Зависимость мембранного
потенциала горизонтальных клеток трех разных типов в сетчатке рыбы от
длины волны монохроматического стимула (ось
абсцисс). Длина волны стимулов с равной энергией изменялась плавно. А. Ответ горизонтальной
клетки–гиперполяризация во всех областях спектра. Б. Красно–зеленая
горизонтальная клетка; зеленый стимул вызывает гиперполяризацию,
красный–деполяризацию. В. Желто–синяя
горизонтальная клетка. На каждом графике черной линией показан уровень
мембранного потенциала покоя. Область под этой линией соответствует
гиперполяризации, над ней–деполяризации (по Spekreijse, Norton: J. gen. Physiol., 1970, 56 с
изменениями)
on– и off–центрами, некоторых ганглиозных клеток и нейронов
ЛКТ. В сетчатке имеются нейроны, активируемые красным и тормозимые зеленым
светом в их РП (простые оппонентные
клетки). Рис. 11.44 демонстрирует дальнейшее развитие принципа оппонентных
цветов применительно к организации рецептивных полей. В данном случае центр и
периферия РП дают противоположные ответы на разные цвета (дважды оппонентные клетки). Нейроны данного типа обнаружены в ЛКТ
и первичной зрительной коре приматов с цветовым зрением. Пример ответов таких
нейронов: если красный свет в центре РП возбуждает их, а зеленый–тормозит, то
на периферии РП картина противоположная. Такая двойная оппонентная организация
РП объясняет феномен одновременного цветового контраста. Обработка
цветоспецифичной информации происходит главным образом в тех участках области
VI, где нет клеток с ориентационно–специфичными РП (рис. 11.28).
Цветоспецифичные нейроны первичной зрительной коры особенно чувствительны к
границам между различно окрашенными областями. Последующая обработка цветовой
информации осуществляется главным образом в корковой области V4, куда идут аксоны из VI, Показано, что V4 включает высокоцветоспецифичные нейроны,
активируемые стимулами со сравнительно узким диапазоном цветовых тонов.
Рецептивные поля некоторых из этих клеток подразделяются на полосы с разной
цветовой спецификой, в результате чего наиболее эффективный стимул для
них–цветовой

Рис. 11.44.
Упрощенная схема пространственной организации двух типов рецептивных полей
нейронов млекопитающих с цветовым зрением (либо ганглиозных клеток сетчатки,
либо нейронов ЛКТ). А. Нейрон
красно–зеленой системы. Б. Нейрон
желто–синей системы. У таких цветоспецифичных РП отмечается антагонизм центра и
периферии. Ц–центр РП; П–периферия РП
контраст. Выводы из этих данных по цветоспецифичным
реакциям нейронов афферентных и центральных отделов зрительной системы приматов
еще обсуждаются, однако в целом намечается следующая картина.
1. Справедливость трехкомпонентной теории на уровне
колбочек несомненна.
2. В сетчатке действует простой механизм оппонентных
цветов.
3. Стадия цветоспецифичной обработки сигналов
нейронами с дважды оппонентным механизмом (рис. 11.44) еще точно не
локализована.
4. Весьма вероятно, что в экстрастриарных зрительных
областях, особенно в V4, происходит более сложная, чем в простой системе оппонентных
цветов, обработка цветовой информации, обеспечивающая относительное постоянство
цветовосприятия [8, 13, 16].
Нарушения цветового зрения
Патологические изменения, нарушающие цветовосприятие,
могут происходить на уровне зрительных пигментов, обработки сигналов в
фоторецепторах, в более центральных отделах нервной системы, а также в самом
диоптрическом аппарате глаза. Очень редко такие нарушения связаны с
повреждением больших полушарий. При «периферических» расстройствах врожденного
характера почти всегда затронуты оба глаза, хотя известны и редкие случаи
типичных нарушений цветового зрения лишь одного из них. В последнем случае
больной сам способен описать субъективное восприятие этих аномалий, поскольку
может сравнить то, что он видит правым и левым глазом. Различают два главных
типа периферического нарушения
цветовосприятия–трихроматический и дихроматический.
Аномалии трихроматического зрения. Самые легкие формы нарушения цветовосприятия обычно наследуются как рецессивный признак,
переносимый Х–хромосомой, поэтому такими аномалиями гораздо чаще страдают
не женщины, а мужчины. Аномальные трихроматы различают меньше цветовых
валентностей, чем люди с нормальным зрением. Как и последние, для описания
видимых цветов они используют три главные валентности–по уравнению (13),–однако
весовые коэффициенты a, b и с у них искажены. Различают три категории аномалий
цветовосприятия. Протаномалы и
дейтераномалы не способны различать ненасыщенные красный и зеленый цвета.
При тестировании на аномалоскопе Протаномалы добавляют в смесь больше, чем в
норме, красного, а дейтераномалы –зеленого. В очень редких случаях тританомалии нарушается работа
желто–синей системы.
Дихроматы. Большинство форм дихроматопсии также передается по
наследству как рецессивные признаки, связанные с Х–хромосомой. Дихроматы могут
подбирать все тона, которые видят, с помощью только двух спектральных цветов (по аналогии с уравнением 13), поэтому по
сравнению с трихроматами число
различаемых ими цветовых валентностей гораздо меньше. У пpoтанопов и дейтеранопов нарушена красно–зеленая система, предположительно
из–за недостатка как пигментов в красных и зеленых колбочках, так и связей
между нейронами сетчатки. Максимум кривой фотопической спектральной яркости у
протанопов примерно 520 нм, а у
дейтеранопов 580 нм. Протанопы путают некоторые красные
тона с черным, темно-серым и коричневым, а в некоторых случаях, подобно
дейтеранопам, с зеленым. Для них часть солнечного спектра в интервале между 480
и 490 нм выглядит ахроматической. Для дейтеранопов этот диапазон составляет
495–500 нм. Очень редко встречающиеся тританопы путают желтый и синий цвета.
Сине–фиолетовый участок спектра кажется им переходом от серого к черному;
область между 565 и 575 для них также ахроматическая. Как
правило, и у дихроматов, и у аномальных трихроматов скотопическое зрение
остается нормальным.
Корковые нарушения цветового зрения. Обширное поражение экстрастриарной зрительной коры
(зона V4) может также привести к нарушению цветовосприятия.
Такие больные неправильно классифицируют цветные карточки, т.е., по–видимому,
затронута его категориальная организация. Подобные корковые нарушения нужно
отличать от затрудненного называния цветов
(цветовая аномия) у больных с патологиями угловой и огибающей извилин левого полушария.
Полная цветовая слепота. Менее 0,01% всех людей являются полностью
цветослепыми. Эти монохроматы видят
окружающий мир черно–белым, т. е. различают лишь оттенки серого цвета. У них
обычно нарушена световая адаптация в фотопических условиях. Их глаза легко
ослепляются даже относительно несильным светом, поэтому они очень плохо
различают форму при дневном освещении, что приводит к фотофобии. Даже в обычный день они носят темные солнцезащитные
очки. Их острота зрения в области центральной ямки снижена до менее 0,1 (угл.
мин)–1. При гистологических

Рис. 11.45.
Пример рисунка из набора «псевдоизохроматических» таблиц Исихары. Испытуемый с
нормальным цветовым зрением видит число «26», протаноп цифру «6»,
дейтераноп–цифру «2»
исследованиях сетчатки монохроматов никаких изменений
обычно не находят. Поскольку кривая спектральной яркости в скотопических
условиях у них нормальна (рис. 11.4), считается, что в их колбочках в качестве
зрительного пигмента содержится родопсин. Об этом же свидетельствуют и их
кривые темновой адаптации (рис. 11.24).
Нарушения палочковой системы. Люди с аномальной палочковой системой воспринимают
цвета нормально, однако их способность к темновой адаптации сильно ограничена.
Причиной такой «куриной слепоты» (никталопии)
может быть недостаток в пище витамина A,
предшественника ретиналя.
Диагностика нарушений цветового зрения. Поскольку нарушения цветового зрения передаются по
наследству как признаки, сцепленные с Х–хромосомой, они гораздо чаще
встречаются у мужчин, чем у женщин. Примерно 0,9% мужчин–Протаномалы,
1,1%–протанопы, 3–4%–дейтераномалы и 1,5%–дейтеранопы. Тританомалия и
тританопия крайне редки. Дейтераномальны примерно 0,3% женщин, протаномальны
0,005%.
Поскольку нормальное цветовое зрение необходимо для
целого ряда профессий (например, летчиков, машинистов, дизайнеров), у всех
детей следует проверять его состояние до того, как они выберут работу и начнут
ей обучаться. В одном из простейших тестов такого рода используются
«псевдоизохроматические» таблицы Исихары (рис. 11.45). На них изображены пятна
разного размера и цвета, расположенные так, что образуют буквы или цифры,
причем у пятен разного цвета одинаковые уровни
светлоты. Лица с нарушенным цветовым зрением не способны видеть некоторые
символы; это зависит от цвета образующих их пятен. Используя различные
комбинации тонов, с помощью таблиц Исихары можно достаточно надежно определить
вид нарушения. Для точной диагностики применяют тесты на смешение цветов,
основанные на уравнениях (12)–(14).
11.8. Литература
Учебники и руководства
1. BakerR., Berthoz A. (eds.). Control of gaze by
brain stem neurons, Amsterdam–New York, Elsevier, 1977.
2. Bing R., Bruckner R. Gehirn und Auge. Grundriss
der Ophthalmo–Neurologie, 3rd Ed. Basel, Schwabe, 1954.
3. Cajal R. S. Die Retina der Wirbeltiere,
Wiesbaden, Bergmann, 1894.
4. Darlnall H. J. A. (ed.). Photochemistry of
vision. Handbook of Sensory Physiology, Bd. VII/1, Berlin–Heidelberg–New York,
Springer, 1972.
5. Davson V. The Eye. 4 vols. London, Academic Press, 1962.
6. Dichgans
J.. Bizzi E. (eds.). Cerebral control of eye movements and motion
perception. Basel, Kargcr, 1972.
7. Drujan B. D. S., Laufer M. (eds.). The S–potentials, New
York, Liss, 1982.
8. Fuorter M. G. F. (eds.). Physiology of
photoreceptor organs. Handbook of Sensory Physiology, Vol. VII/2. Berlin
Heidelberg–New York, Springer, 1972.
9. Graham С. H. (ed.). Vision and
visual reception. New York–London, Sidney, J. Wiley, 1965.
10. Granil R. Receptors and sensory perception. New Haven, Yale
University Press, 1955.
11. Grusser O.–J., Klinke R. (eds.). Zeichnerkennung durch biologische
und technische Systeme. Berlin–Heidelberg–New York, Springer, 1971.
12. Helmholtz H. von: Handbuch der Physiologischen Optik, 2nd Ed.
Hamburg, Leipzig, L. Voss, 1896.
13. Hering E. Grundzuge der Lehre vom Lichtsinn, Berlin, Springer,
1920.
14. Hofmann F.B. Die Lehre vom Raumsinn des Auges (1920).
Berlin–Heidelberg New York, Springer, 1970 (Reprint).
15. Hyv'drinen J. The parietal cortex of monkey and man.
Berlin–Heidelberg–New York, Springer, 1982.
16. Jameson D., HwvichL.M. (eds.). Visual Psychophysics. Handbook of
Sensory Psychology, Vol. VII/4, Berlin–Heidelberg–New York, Springer, 1972.
17. Jung R. (ed.). Central processing of visual information A. Integrative
function and comparative data. Handbook of Sensory Psychology, Vol. VI1/3A,
Berlin – Heidelberg, New York, Springer, 1973.
18. Jung R. (ed.). Central processing of visual information. B.
Visual centers in the brain.
Handbook of Sensory Physiology. Vol. VII/3B, Berlin–Heidelberg–New York,
Springer, 1973.
19. Kommereli G. (ed.). Augenbewegungsstorungen. Neurophysiologie und
Klinik, Munchen, J. F. Bergmann, 1978.
20. Landolt E. Die Untersuchung der Refraction und der Akkommodation.
In: Graefe–Saemisch's Handbuch der gesamten Augenheilkunde, 3rd Ed.,
Untersuchungsmetho–den, Vol. 1, Berlin, Springer, 1930.
21. Lennerstrand G., Bach–Y–Rita P. Basic mechanisms of ocular motility
and their clinical implications. Oxfomd, Pergamon Press, 1975.
22. Linksz A. Physiology of the eye. Vol. I, Optics. Vol. II, Vision,
New York, Gruner und Stratton, 1950–1952.
23. Mitize К., Nehrling В., Rentier J. Brillenglasbestimmung. Zurich
Verlag fur Augenheilkunde und Optik, 1972.
24. Polyak S. The vertebrate visual system, Chicago, University of
Chicago Press, 1957.
25. Schober H. Das Sehen, Vol. 2, 2nd Ed. Leipzig–Fachbuch–verlag,
1958.
26. Siebeck R. Optik des menschlichen Auges,
Berlin–Gottingen–Heidelberg, Springer, 1960.
27. Spieyel I. M. (ed.). Readings in the study of visually perceived
movement. New York, Harper and Row, 1965.
28. Stohr M., Dichgans J., Diener H. C., Buettner U. W. Evozicrte
Potentiate, Berlin – Heidelberg – New York, Springer, 1982.
29. Walls G. L. The vertebrate eye and its adaptive radiation, New York,
London, Hafner, 1963.
30. Walsh F.B.. Hoyt W.F. Clinical neuroophthalmology, 3rd Ed.
Baltimore, William & Wilkins, 1969.
31. Wright W.D. The measurement of colour, 3rd Ed. London, Hilger und
Watts, 1964.
32. Yarhus A. L. Eye movements and vision. New York, Plenum Press,
1967.
33. Zrenner E. Neurophysiological aspects of colour vision in primates,
Berlin – Heidelberg – New York, Springer, 1983.
Оригинальные статьи и обзоры
34. Altman J. New visions in photoreception. Nature, 313, 264–265
(1985).
35. Baumgartner G., Hakas P. Die Neurophysiologie des simultanen
Helligkeitskontrastes, Pflug. Arch. ges. Physiol., 274, 489 (1962).
36. Baylor D. A., Fuortes M. G. F. Electrical responses of single cones
in the retina of the turtle. J. Physiol. (Lond.), 207, 77 (1970).
37. Bill A. Uveoscleral drainage of aqueous humour in human eyes. Exp.
Eye Res., 12, 275 (1971).
38. Boycott В. В., Wassle H. The morphological types of
ganglion cells of the domestic cats' retina. J. Physiol. (Lond.), 240, 397
(1974).
39. Coliewijn H. M., Curio G., Grusser O.–J. Spatially selective visual
attention and generation of eye pursuit movement. Experiments with Sigma
movement. Human Neurobiology, 1, 129 (1982).
40. Desimone R., Alhrighl Т.О., Gross С.,
Bruce С. Stimulus selective properties
of inferior temporal neurons in the macaque, J. Neurosciences, 4, 2051 (1984).
41. Die G. van, Coliewijn H. Optokinetic nystagmus in man. Role of
central and peripheral retina and occurrence of asymmetries. Human Neirobiol.,
1, 111 (1982).
42. Essen D. C. van. Visual areas of the mammalian cerebral cortex.
Ann. Rev. Neurosciences, 2, 227 (1979).
43. Essen D. C. van, Mounsell J. H. R.., Bixby J. L. The middle
temporal visual area in the macaque: myeloarchitecture connections, functional
properties and topographic organization, J. Сотр. NeuroL, 199, 293
(1981).
44. Essen D. C. van, Newsome W. Т.,
Bixby J. L. The pattern of intrahemispheric connections and its relationship to
extrastriate visual areas in the macaque monkey, J. Neuroscience 2, 265 (1982).
45. Essen D. C. van, Zekl S. M. The topographic organization of Rhesus
monkey prestriate cortex, J. Physiol., 277, 193 (1978).
46. Fesenko E. E., Kolesnikov S. S.. Lybarsky A. L. Induction by cyclic
GMP of cationic conductance in plasma membrane of retinal rod outer segment.
Nature, 313, 310 (1985).
47. Grusser O.–J. Grundlagen der neuronalen Infonnations–verarbeitung
in den Sinnesorganen und im Gehirn. Informatik–Fachberichte Bd. 16, 34.
Berlin–Heidelberg–New York, Springer, 1978.
48. Grusser O.–J. Multimodal structure of
the extrapersonal space. In: Hem A.,
Jeannerod M. Spatially oriented behaviour. New York–Berlin–Heidelberg–Tokyo, Springer, 1982, р. 328.
49. Grusser O.–J. Die functionelle Organisation der Saugetiernetzhaut–physiologische
und pathophysiologische Aspete, Fortschr. Ophthalmol., 80, 502 (1983).
50. Grusser O.–J. Face recognition within the region of neurobiology
and beyond it. Human Neurobiol., 3, 183 (1984).
51. Grusser O.–J., Grusser–Cornehls U. Periodische Aktivierungsphasen
visueller Neurone nach kurzen Lichtreizen verschiedener Dauer. Pflug. Arch.
ges. Physiol., 275, 292 (1962).
52. Grusser O.–J., Grusser–Cornehls U. Neurophysiologie des
Bewegungssehens. Bewegungsempfindliche und richtungs–spezifische Neurone im
visuellen System. Ergebn., Physiol., 61, 178 (1969).
53. Hagins W. A., Penn R.D., Yoshikami S. Dark current and photocurrent
in retinal rods. Biophys. J., 10, 380 (1970).
53а. Непп V.,
Buttner–Ennever J. A., Hepp К. The primate oculomotor
system I and II, Human Neurobiol., 1, 77 and 87 (1982).
54. Hubel D. H., Wiesel Т. N. Receptive fields
and functional architecture of monkey striate cortex, J. Physiol. (Lond.), 195,
215 (1968).
55. Hubel D. H., Wiesel T. N. Cell sensitive to binocular depth in area
18 of the macaque monkey cortex. Nature, 225, 41 (1970).
56. Hubel D.H., Wiesel T.N. Functional architecture of macaque visual
cortex, Proc. Roy. Soc. (Lond.) B, 198, 1 (1977). 56a. Lamb T.D. Transduction in vertebrate photoreceptors: the roles of
cyclic GMP and calcium, Trends in Neurosciences, 9, 224 (1986).
57. Livingston M. S., Hubel D. H. Anatomy and physiology of a color
system in the primate visual cortex, J. Neurosci., 4, 309 (1984).
58. Lynch J. C. The functional organization of posterior parietal
association cortex. Behavior and Brain Science, 3, 485 (1980).
59. Lynch J.C., Mountcastle V.B„ Talbot W.H., Yin 7: С. Т Parietal lobe
mechanisms for directed visual attention, J. Neurophysiol, 40, 362 (1977).
59a. Perett D.I., Rolls E. T, Kaan
W. Visual neurons responsive to faces in the monkey temporal cortex. Exp.
Brain Res 47, 329 (1982).
60. Schiller P. H. The role of the monkey superior colliculi in eye
movement and vision. Invest. Ophthal, 11, 451 (1972).
61. Tomita T. Electrical activity of vertebrate photoreceptors. Quart.
Rev. Biophys., 3, 179 (1970).
62. WildH.M., Butler S.R., Garden Kulikowski J.J. Primate cortical area
V4 important for colour constancy but not wavelength discrimination, Nature,
313, 133 (1985).
63. Wurtz R. H., Goldberg M. E. The primate superior colliculus and the
shift of visual attention. Invest. Ophthal. 11, 441 (1972).
64. Zeki S. M. Functional specialization in the visual cortex of Rhesus
monkey, Nature, 274, 423 (1978).
65. Zeki S. M. Uniformity and diversity of structure and function in
Rhesus monkey prestriate visual cortex, J. Physiol. (Lond.), 277, 273
(1978).