ОБЩАЯ СЕНСОРНАЯ ФИЗИОЛОГИЯ
X. Хандверкер
8.1.
Предмет изучения общей сенсорной физиологии
Та ветвь сенсорной физиологии, которая называется «общей»,
изучает общие принципы, лежащие в основе сенсорных способностей, т.е. работы
отдельных сенсорных систем и их
результата–субъективного восприятия
человека. Два этих аспекта требуют разных стратегий исследования. Мы говорим об
объективной сенсорной физиологии, когда анализируются физические и химические
параметры работы сенсорных систем. Если же психологическими методами изучается
субъективное восприятие человека, а полученные результаты используются для
описания сенсорных функций, речь идет о субъективной сенсорной физиологии.
Объективная и субъективная сенсорная физиология
Из всего множества факторов окружающей среды,
воздействующих на наш организм, некоторые (далеко не все) улавливаются
сенсорными органами. Такие факторы называют
сенсорными стимулами. Под их влиянием рецепторные клетки генерируют
потенциалы, которые активируют афферентные нервные волокна. Импульсация многих
афферентов проводится к сенсорным центрам в мозгу, где производится обработка
информации. Эту цепь физико–химических событий можно исследовать методами,
используемыми и в случае других физиологических процессов. Здесь речь идет об объективной сенсорной физиологии.
Поскольку сенсорные органы человека и животных устроены сходным образом, можно
сформулировать некоторые ее общие правила.
Сенсорный стимул часто приводит к возникновению субъективного ощущения. Например, электромагнитные колебания с
длиной волны 400 нм вызывают ощущение «голубого цвета».
Описывая его, человек говорит: «Я вижу голубое
пространство». Обычно мы сразу же пытаемся интерпретировать наши ощущения на
основе предшествовавшего опыта, т.е. преобразуем их в восприятие. Ощущение «Я вижу голубое пространство» может
соответствовать восприятию «небо». Влияние опыта на восприятие может
сказываться еще сильнее: метеоролог увидит в небе не просто белые пятна, а
слоисто–кучевые облака, иллюстратор детских книг–овец на синей лужайке. На
восприятие могут влиять и многие психологические факторы, например настроение.
Свое восприятие мы осознаем
непосредственно (без помощи измерительных приборов), т. е. совершенно иным
образом, нежели рецепторные потенциалы и потенциалы действия, регистрируемые
методами объективной сенсорной физиологии. Пытаясь на основе субъективных
ощущений и восприятии разобраться в работе сенсорных органов, мы вступаем в
область субъективной сенсорной
физиологии.
«Загадочная картинка» (рис. 8.1)–хорошая иллюстрация
того факта, что восприятие не просто фотографическое отображение окружающего
сенсорными органами. Изображенный объект может восприниматься как голова либо
зайца, либо утки. Если смотреть на картинку некоторое время, два эти восприятия
будут чередоваться, хотя никаких изменений в информации, поступающей от глаз,
не происходит. Кроме того, мы не можем одновременно увидеть и зайца, и утку,
даже зная, что изображение «двузначно». Очевидно, у восприятия есть
определенная структура, обеспечивающая однозначную интерпретацию увиденного с
помощью активных интегративных процессов, протекающих в мозгу [6]. Начиная с
прошлого века, физиологи, изучавшие сенсорные органы, старались главным образом
установить количественные соотношения между интенсивностью стимула и силой
субъективного ощущения. Этот подход, называемый психофизикой, применяется как в сенсорной

Рис. 8.1.
«Зайцеутка»– двузначное изображение (автор–Jastrow), опубликованное впервые
примерно в 1900г. (из [6])
физиологии, так и в экспериментальной психологии, или,
точнее говоря,–в психологии восприятия. В
последнее время, однако, появилась возможность объединить измерение
физиологических характеристик сенсорных органов и возникающих при этом
ощущений, т.е. непосредственно сравнивать их между собой. Подобные исследования
относят к области психофизиологии, или
интегративной сенсорной физиологии.
На рис. 8.2 показана упрощенная схема отображения каждого
из сенсорных уровней на следующем. Черные стрелки означают «вызывает» или
«приводит к». Пунктирная красная стрелка демонстрирует переход от физических
параметров к психическим. Можно ли на этом уровне интерпретировать выражение
«приводит к...» как причинно–следственную связь?
Связь психического и физического. Веками философы строили различные теории относительно
природы сознания и его взаимоотношений с телом. В настоящее время доминируют
две основные концепции. Согласно одной из них, сложные процессы, идущие в мозгу
в ответ на предъявления сенсорного стимула, идентичны субъективным ощущениям.
Два эти аспекта просто разные способы рассмотрения одного и того же
явления–функционирования мозга.
Это–монистическая точка зрения. Согласно ей пунктирная стрелка на рис. 8.2
вообще не отражает какой–либо связи и должна быть заменена знаком равенства.
С другой точки зрения, дуалистической, мозг–сложный инструмент независимого от него
сознания (или души). В этом случае пунктирная стрелка на рис.
8.2–причинно–следственная связь: мозговые процессы воздействуют на сознание и
наоборот.
Как осуществляется такое взаимодействие, вне нашего
понимания. Проблема связи психического (сознание) и физического
(тело)–философская и не решается методами естественных наук [II]. Это следует
из нашей неспособности поставить эксперимент, который бы опровергал (в смысле
Попера [22]) либо монистическую, либо дуалистическую точку зрения.
Нейрофизиологи и психологи также отдали дань философскому осмыслению проблемы
связи физического и психического [12, 23].
Парапсихология.
Не может ли .решить эту проблему парапсихология? Она занимается эмпирическим
изучением таких феноменов, как телепатия и ясновидение, объединяемых термином экстрасенсорное восприятие (ЭСВ).
Делались попытки различным способом продемонстрировать их экспериментально.
Если бы реальность ЭСВ подтвердилась, это был бы, разумеется, сильный аргумент
в пользу существования независимого от тела духа, обладающего экстрасенсорной
информацией. Тем не менее не все дуалисты верят в ЭСВ.
Пока парапсихологические исследования не дали
убедительных для скептиков доказательств ЭСВ [14]. Трудность заключается,
вероятно, не в том, что парапсихологи как исследователи не признаны «научными
авторитетами» (как они сами считают), а скорее в неадекватности используемых
«обычных» методов для анализа феноменов ЭСВ. Здесь две основные причины.
Во–первых. такие феномены, если и существуют, наблюдаются крайне редко и в
условиях, которые трудно проконтролировать, т.е. они невоспроизводимы. При этом
часто делаются ошибки при статистической интерпретации результатов.

Рис. 8.2. Схема взаимоотношений в сенсорной физиологии. Прямоугольниками обозначены основные
рассматриваемые ею феномены, стрелки
между ними соответствуют связям типа «приводит к» или «вызывает» (при
условиях.–перечисленных под каждой стрелкой). Пунктирная стрелка–это переход от физиологических процессов к
психическим
Например, если игрок в кости выбросил три раза подряд
шестерку, считается, что он удачно сыграл, а вовсе не продемонстрировал способность
к ЭСВ. И нет смысла задним числом вычислять вероятность этого события (она
составляет всего 1/6 х 1/6 х 1/6, т.е. менее 0,5%), поскольку оно уже произошло
«на 100%». Оно доказало бы необычные способности игрока, только если бы он
заранее сказал, что выбросит подряд три шестерки. Даже в этом случае следовало
бы сначала проверить, нет ли здесь простого фокуса. Вот и вторая причина
трудностей с подтверждением ЭСВ: история парапсихологии полна обманов, на
которые поддавались даже критически настроенные ученые [14]. Лица со
«способностями» к ЭСВ часто оправдывают свое желание уклониться от строгих
экспериментальных условий тем, что такая обстановка на них плохо влияет. Тем не
менее эксперименты в этой области требуют очень точного контроля, умения обнаруживать
трюки и досконального знания приемов профессиональных фокусников. Такое нечасто
встретишь среди ученых.
Самостоятельная ценность физиологических
и психологических правил. Может
показаться, что монистический взгляд на связь физического и психического делает
излишним разделение сенсорной физиологии на субъективную и объективную. Однако
это не так. Независимо от того, «идентичны» психические процессы мозговым
функциям или нет, у каждой реальности–и физической, и психической–есть свои
собственные правила, концепции и терминология. Их самостоятельность можно
проиллюстрировать на примере компьютера. Исчерпывающее знание его устройства
само по себе не гарантирует понимания функциональных законов языков
программирования, а тем более содержания конкретных программ. Так и
исчерпывающее знание анатомии и физиологии головного мозга не помогает
объяснить человеческое поведение. Тем не менее, как подробно обсуждается в
разд. 8.4, посвященном «интегративной сенсорной физиологии», психика
определяется именно его работой. Именно объективная сенсорная физиология
позволяет выяснить границы нашего восприятия. Так и знание компьютера
необходимо, чтобы определить максимальный объем его программ, скорость их
выполнения, входные переменные и возможные периферические устройства.
8.2.
Общая объективная сенсорная физиология
Специфичность сенсорных органов
В ходе эволюции у всех организмов развились
специализированные сенсорные органы, устроенные так, чтобы оптимальным образом
отвечать на вполне определенные стимулы. Обычно, наблюдая за реакциями
организма, легко определить, какой стимул для данного сенсорного органа
оптимален. Как правило, он возбуждает его минимальным
количеством энергии. Адекватный стимул для данного органа–тип раздражения, вызывающий
оптимальный ответ. Например, палочки и колбочки сетчатки глаза можно возбудить,
сильно надавив на глазное яблоко; при этом возникают зрительные ощущения.
Однако оптимальные (и, следовательно, адекватные) стимулы в данном
случае–электромагнитные колебания с длиной волны от 400 до 800 нм. Чем выше
специфичность сенсорного органа, тем больше вероятность его возбуждения (при
физиологических условиях) только адекватными стимулами.
Структура сенсорных органов как причина
их специфичности. Во многих
случаях адекватность стимула определяется не только свойствами рецепторных
клеток, но и макроструктурой органа. Например, адекватный стимул для рецепторов
и вестибулярного аппарата, и внутреннего уха (в обоих случаях это волосковые
клетки)–поток эндолимфы, отклоняющий реснички клеток. Однако структура
внутреннего уха такова, что эндолимфа приходит в движение, когда механические
колебания частотой от 20 до 20 000 Гц достигают улитки, в то время как в
вестибулярном аппарате эндолимфа смещается при изменении положения головы.
Реакции сенсорных органов на разные виды
стимулов. Если
сравнивать разные химические и физические стимулы, не всегда просто определить,
чисто формально учитывая только энергию воздействия, какой из них для данного
сенсорного органа адекватен. Например, холодовые рецепторы в слизистой рта и
носа реагируют не только на охлаждение, но и весьма чувствительны к
определенным химическим стимулам, в частности ментолу. Если курить ментоловую
сигарету, их возбуждение вызывает ощущение холодного дыма. Другой
пример–тепловые рецепторы кожи. Возбуждаясь при повышении температуры, они
также весьма чувствительны к увеличению концентрации Са2+ во
внеклеточном пространстве. Это происходит, например, после инъекции в артерию
раствора, содержащего кальций. В результате возникает ощущение тепла в обслуживаемом
ею участке тела.
Закон «специфических сенсорных энергий», сформулированный 150 лет
назад Иоганнесом Мюллером, гласит, что характер ощущения определяется не
стимулом, а раздражаемым сенсорным органом. Приведенные выше примеры доказывают
его справедливость. Это один из важнейших законов субъективной сенсорной
физиологии.
С точки зрения объективной сенсорной физиологии
специфичность органа определяется характером обработки возникающих в нем
сигналов центральными отделами нервной системы. Например, кожные тепловые
рецепторы ответственны за терморецепцию и терморегуляцию главным образом
благодаря своим центральным соединениям.
Категории ощущений.
Сенсорные органы можно подразделить на три группы следующим образом.
1. Первое, что ассоциируется со словом «ощущения»,–это
органы и рецепторы, стимулируемые окружающей средой. Все они относятся к экстероцепторам.
2. Другие органы определяют длину мышц, натяжение
сухожилий, углы в суставах и другие параметры положения и движений тела. Их
называют проприоцепторами. К этой
группе можно также отнести вестибулярный аппарат.
3. Наконец, сенсорная информация поступает и от
внутренних органов тела. Идущие от них афференты носят название интероцепторов.
Большая часть информации, посылаемой в ЦНС
интероцепторами и проприоцепторами, очень редко доходит до нашего сознания или
вообще никогда им не воспринимается. Например, мы и не подозреваем о сигналах
от барорецепторов, расположенных в
каротидном синусе, которые непрерывно контролируют артериальное давление крови.
Долгое время не было ясно, доходят ли до сознания импульсы от мышечных веретен.
Ответ на этот вопрос в конце концов получил Маклоски [20], поставив эксперимент
на самом себе. Обнаженная хирургическим путем мышца раздражалась стимулами,
адекватными для первичных (Ia) афферентов
мышечных веретен. Полученный результат однозначно свидетельствовал, что
увеличение частоты импульсации этих нервных волокон ощущается как изменение
положения соответствующей конечности.
Процесс преобразования (трансдукции)
В каждом сенсорном органе есть «рецепторы», с
возбуждения которых начинается сенсорный процесс. К сожалению, терминология в
последнее время стала неоднозначной. Исходно под рецептором понимали сенсорную
клетку, однако сейчас так называют в молекулярной биологии комплексы молекул на
клеточных мембранах, специфически реагирующие с другими молекулами, например
гормонами. Даже в сенсорной физиологии термин «рецептор» используется
неоднозначно. Анатомы считают рецептором морфологически цельную сенсорную
клетку, а физиологи–часть мембраны такой клетки или нервное окончание,
специализированные для восприятия стимулов. Мы будем придерживаться
«компромиссного» толкования и называть рецептором клетку или ее часть, которая
ответственна за преобразование стимулов в нейронное возбуждение. Часто
рецепторами служат окончания периферических аксонов или дендритов афферентных
нервных волокон. С другой стороны, в
некоторых сенсорных органах такие окончания соединены
со специализированными сенсорными клетками, не являющимися нейронами (например,
волосковыми клетками в улитке). Наконец, в сетчатке глаза особые сенсорные
клетки (палочки и колбочки) имеют нейронное происхождение. Все это также
рецепторы.
Рецепторный потенциал. Рецепторы преобразуют энергию стимула в изменение
проницаемости своей мембраны. Этот процесс называют трансдукцией. Вместе с проницаемостью меняется и потенциал
мембраны: он становится «рецепторным». Поскольку рецепторный потенциал
генерирует в афферентных нервных волокнах потенциалы действия, его также
называют генераторным.
Определение рецептора. Данное выше определение можно теперь сформулировать
иначе, используя представление о потенциалах: рецептор–это клетка или часть ее
мембраны, генерирующая рецепторные потенциалы, которые кодируются и передаются
по соответствующим афферентам в виде последовательностей потенциалов действия.
В известных рецепторах кожи, внутренних органов и мышц рецепторные потенциалы
возникают в самих окончаниях афферентов, т.е. трансдукция происходит
непосредственно в нервных окончаниях. Последние либо свободно лежат в тканях,
либо заключены в особые структуры типа «тельца» или мышечного веретена.
Изменения потенциала, возникающие при стимуляции
рецептора, можно записать с помощью внутриклеточных микроэлектродов. Три
примера измерения генераторного потенциала показаны на рис. 8.3. В двух случаях
(мышечное веретено и тельце Пачини) рецептором служит окончание аксона, а в
третьем–особая волосковая клетка. Считается, что в последнем случае рецепторный
потенциал возбуждает афферентный аксон путем синаптической передачи, медиатор
которой еще не известен (см. также гл. 12).
В большинстве случаев молекулярные механизмы
трансдукции стимула в рецепторный потенциал мембраны выяснены недостаточно.
Одна из причин этого–обычно очень малый размер и часто относительная недоступность
рецептивных участков клеток. В принципе трансдукция включает открывание каналов
мембраны, что позволяет проходить через нее потоку ионов, обусловленному
разницей их концентраций внутри и вне клетки. Мембранными каналами управляют
особые молекулярные комплексы, которые меняются под действием стимула.
Например, в хеморецепторах связывание молекул рецептивной мембраны с адекватным
веществом влияет на молекулу, открывающую ионные каналы. На их размеры в случае
механорецепторов может

Рис. 8.3. Различные
типы рецепторов. А. Волосковая клетка
в улитке или вестибулярном органе. Б.
Мышечное веретено лягушки. В. Тельце
Пачини. Звездочкой отмечены
предполагаемые места трансформации генераторного потенциала в потенциал
действия (по Flock в [2] с изменениями)
воздействовать растяжение мембраны. Рецепторные
потенциалы во всех этих примерах означают деполяризацию. В фоторецепторах
сетчатки (палочках и колбочках) ионные токи возникают главным образом в темноте
и блокируются при попадании света на мембранные диски наружных сегментов
клеток. В данном случае рецепторный потенциал соответствует гиперполяризации.
Свойства рецепторных потенциалов в афферентных нервных
окончаниях следующие.
1. Они генерируются в самих нервных окончаниях, а не в окружающих клетках, входящих в
структуру сенсорного органа. Это хорошо видно на примере тельца Пачини (рис.
8.4), аксон которого возбуждается механическими стимулами даже тогда, когда
окружающая его луковицеобразная капсула удалена. В более проксимальной части
клетки, где генерируются и проводятся потенциалы действия, аксонная мембрана
гораздо менее чувствительна к механическому раздражению.
Равновесный потенциал мембраны тельца Пачини –
примерно 0 мВ. Значит, изменения проницаемости, вызываемые стимулом, не
ограничиваются

Рис. 8.4.
Генераторные потенциалы и потенциалы действия тельца Пачини. 1. Слабое
раздражение изолированного рецептора в месте, отмеченном стрелкой, вызывает рецепторные (генераторные) потенциалы, амплитуда
которых кодирует интенсивность стимула (а–г). Надпороговые рецепторные
потенциалы запускают потенциалы действия (д). 2. Даже после устранения слоев,
окружающих рецептор, при его механической стимуляции возникают генераторные
потенциалы. 3. Давление, приложенное к первому перехвату Ранвье афферентного аксона,
блокирует генерирование потенциалов действия, но не влияет на рецепторный
потенциал. Внизу: запуск потенциалов
действия в первом перехвате Ранвье за счет электротонического распространения
генераторного потенциала (по [19] с изменениями)
натриевыми каналами. Запись активности этих рецепторов
in vitro
показала, что при удалении из окружающего пространства всех ионов Na+
рецепторный потенциал уменьшается примерно до 1/3 контрольного значения, но не
исчезает совсем. Кроме того, как известно, каналы, участвующие в генерировании
рецепторного потенциала, почти не чувствительны к тетродотоксину (ТТК), чем
сильно отличаются от натриевых каналов в аксонных перехватах Ранвье,
ответственных за распространение потенциала действия. Тельца Пачини продолжают
генерировать рецепторные потенциалы даже в тех случаях, когда проведение
потенциалов действия в афферентном волокне блокировано ТТК.
2. Рецепторный потенциал – градуальный. Стимулами разной интенсивности большинство рецепторов
деполяризуется (или гиперполяризуется, как в случае палочек и колбочек)
неодинаково. Хотя амплитуда потенциала определенным образом отражает силу
стимуляции, последняя не служит источником энергии для такого изменения клетки.
Как говорилось выше, единственная функция стимула – управление ионными
токами через мембрану.
Рецептивные мембраны многих рецепторов чрезвычайно
чувствительны к адекватным для них стимулам. По–видимому, в некоторых случаях
их чувствительность достигает теоретически возможного предела. Например,
волосковые клетки улитки возбуждаются при смещении на расстояние, сравнимое с
диаметром атома водорода. В сетчатке достаточно одного фотона на палочку, чтобы
возникли ионный ток через ее мембрану и генераторный потенциал, оказывающий
заметное воздействие на активность ганглиозных клеток, связанных с данной
палочкой. Другими словами, трансдукция включает и процесс усиления.
3. Рецепторный потенциал–локальный; он
распространяется по мембране электротонически, а не проводится активно.
4. Рецепторные потенциалы могут подвергаться пространственной и временной суммации.
Например, если одновременно на тельце Пачини действуют два слишком слабых
поодиночке стимула, вместе они способны вызвать надпороговую деполяризацию. Она
может наблюдаться и в том случае, когда два слабых стимула так быстро следуют
друг за другом, что второй рецепторный потенциал суммируется с первым.
Таким образом, у рецепторных потенциалов много общих
свойств с локальными синаптическими токами центральных нейронов.
Трансформация генераторных потенциалов в
залпы потенциалов действия
На следующем этапе возбуждения рецепторов генераторный
(рецепторный) потенциал приводит к возникновению последовательности потенциалов
действия. Эта трансформация обычно происходит на первом перехвате Ранвье
афферентного нервного волокна. Некоторые афференты не миелинизированы
(например, ноцицепторы и «тепловые» волокна), и точное место трансформации в
них неизвестно. Генераторный потенциал должен распространяться электротонически
до места генерирования потенциала действия точно так же, как синаптический
потенциал по телу мотонейрона к аксонному холмику. У некоторых сенсорных клеток
(например, волосковых во внутреннем ухе и
фоторецепторов сетчатки) между рецепторными потенциалами и потенциалами
действия, вероятно, вклиниваются синаптические процессы. «Генераторными» в
строгом смысле слова здесь будут постсинаптические потенциалы, например,
ганглиозных клеток сетчатки.
На рис. 8.5 в качестве примера изображены потенциалы
действия и генераторные потенциалы мышечного веретена лягушки. Если у генераторного
потенциала амплитуда деполяризации (у фоторецепторов гиперполяризации) отражает
интенсивность стимула, у распространяющихся потенциалов действия она
подчиняется закону «все или ничего», и интенсивность стимула кодируется их
частотой. Частота импульсации в афферентном нервном волокне пропорциональна
величине генераторного потенциала (см. также рис. 8.2). Такое же перекодирование локального потенциала
с переменной амплитудой в проводимый
сигнал с переменной частотой
происходит еще раз в синапсах ЦНС.
Временной аспект сенсорного возбуждения:
фазические и тонические ответы, адаптация
Как видно из рис. 8.5, ответ афферентных волокон
мышечного веретена лишь приблизительно воспроизводит временной ход стимуляции.
Рецептор и его афферент сильнее реагируют на быстрое нарастание интенсивности
стимула, сигнализируя о скорости его изменения. Это называют динамическим, фазическим или дифференциальным ответом. Последний
термин используется по той причине, что скорость изменения (V)–это первая производная расстояния по времени: dL/dt = V.
С другой стороны, если ответ рецептора мало зависит от
скорости изменения стимула, он называется
тоническим, статическим или
пропорциональным. В
большинстве случаев даже тоническая реакция при длительном действии неизменного
стимула медленно снижается: происходит адаптация рецептора. Изображенный на
рис. 8.5 рецептор обладает как дифференциальными, так и пропорциональными
характеристиками (ПД–тип). Разные рецепторы и афференты значительно различаются
по своей фазической и тонической чувствительности.
Причины различий в скорости адаптации
разных рецепторов. Тельце
Пачини работает как детектор ускорения. Это связано с его чрезвычайно быстрой
адаптацией, которая объясняется характерной слоистой природой окружающей
нервное окончание структуры. Ее крайне компактное строение действует как
высокочастотный фильтр, экранирующий рецептор от непрерывного давления и
пропускающий только высокочастотную составляющую механической стимуляции.

Рис. 8.5. Генераторные
потенциалы и серии потенциалов действия в мышечном веретене лягушки и влияние
на них тетродотоксина. Вверху: запись
от ствола афферентного аксона вблизи окончаний сенсорного нерва (А) и в более
проксимальной части (Б) при разных
величинах растяжения мышцы (а, б, в; временной ход стимуляции–внизу слева). Черными кривыми показаны
потенциалы действия; они либо накладываются на рецепторные потенциалы (А), либо
(6) при регистрации дальше от рецептора существуют самостоятельно. Красные кривые – записи после блокады
потенциалов действия тетродотоксином. Скорость деполяризации и частота
потенциалов действия зависят от амплитуды деполяризации рецепторного
потенциала. Внизу: схема средней
части мышечного веретена лягушки с участками отведении А и Б (из Ottoson, Shepherd в [2])
Однако это не единственная причина различий рецепторов
по скорости адаптации, причем такие различия не всегда связаны со структурными
особенностями. Скорость адаптации определяется также процессом трансформации
генераторного потенциала в залп потенциалов действия. Например, в том случае,
когда первый перехват Ранвье тельца Пачини деполяризован длительным
электрическим импульсом, генерируется только один–два потенциала действия. Если
же такой же эксперимент выполняется с первым перехватом Ранвье афферента
мышечного веретена лягушки, длительная деполяризация приводит к длинной серии
потенциалов действия. В конечном счете, скорость адаптации определяется
проводящей мембраной на этапе трансформации генераторного потенциала в
потенциалы действия.
Наиболее распространенный вид адаптации.
При передаче информации в сенсорных системах разные виды
адаптации наблюдаются то чаще, то реже. Хотя некоторые афференты адаптируются
чрезвычайно быстро (например, у телец Пачини), а другие крайне медленно (например,
вторичные афференты мышечных веретен); большинство их относится к ПД–типу (см.
рис. 8.5). Это позволяет передавать в нервную систему информацию о силе
раздражения (пропорциональный компонент ответа), подчеркивая его быстрые
изменения, которые особенно важны для регуляторных процессов (рефлексов),
повышением частоты импульсации (дифференциальный компонент). В большинстве
сенсорных путей такая преимущественная передача информации относительно быстрых
изменений стимуляции сопровождается еще дополнительным усилением в высших
отделах ЦНС.
Пространственный аспект сенсорного
возбуждения; рецептивные поля
Кожные рецепторы возбуждаются стимулами, действующими
на кожу непосредственно над ними. У их афферента бывает несколько ветвей,
каждая из которых иннервирует отдельный рецептор; такое волокно будет
возбуждаться стимулами в пределах более обширной области. Размер его рецептивного поля зависит от степени
ветвления. Мы будем называть такие рецептивные поля первичными в отличие от рецептивных полей центральных нейронов
(см. рис. 8.6).
Разное количество первичных афферентов сходится
(конвергирует) на каждом центральном сенсорном нейроне, рецептивное поле
которого может в результате быть обширнее первичного.
Для большинства сенсорных модальностей известно характерное
пространственное распределение размеров рецептивных полей первичных и высших
сенсорных нейронов. Например, у многих кожных афферентов на кончиках пальцев
рук они меньше, чем на предплечье или туловище. На более высоких уровнях это
различие еще более выражено. В соматосенсорных проекционных областях коры
рецептивные поля нейронов «пальцев» гораздо меньше, чем у нейронов «туловища».
То же самое справедливо и для сетчатки: рецептивные поля ганглиозных клеток,
связанных с рецепторами центральной ямки, меньше, чем у возбуждаемых
рецепторами с периферии сетчатки. В зонах с высокой

Рис. 8.6.
Схема сенсорной системы (по [5] с изменениями)
плотностью иннервации пространственное разрешение стимулов выше (см. также
гл. 9 и 11).
Сенсорные системы в ЦНС: специфичные и
неспецифичные пути
Входя в спинной или головной мозг, первичные афференты
образуют синапсы с вторичными сенсорными нейронами. Аксоны последних идут
вместе в виде сенсорных трактов к ядрам более высоких уровней. Обычно в каждой
сенсорной системе такой восходящий путь проходит через несколько сенсорных
центров. Для всех модальностей, кроме обоняния, к самым высшим из них относятся
ядра таламуса и соответствующие им корковые проекционные зоны. Таким образом,
сенсорный путь состоит из последовательности синаптически связанных друг с
другом центральных нейронов, поочередно возбуждаемых импульсацией
соответствующих рецепторов. Вся совокупность нейронных соединений в пределах
такого пути вместе с тормозными цепями, связанными с ними, образует сенсорную систему.
Схема на рис. 8.6 иллюстрирует некоторые ее
характерные особенности. Первичные афференты обычно ветвятся в сенсорном органе
и иннервируют по нескольку рецепторов, образуя первичное рецептивное поле. Они ветвятся и проксимально,
контактируя синаптически с несколькими сенсорными нейронами: это называется дивергенцией. И напротив, каждый
вторичный нейрон образует синапсы с несколькими первичными афферентами, т. е.
происходит также конвергенция. Такие
же сложные связи присутствуют и в высших сенсорных центрах. Значит, хотя
сенсорные пути можно рассматривать
как последовательности
нейронов, за счет конвергенции и дивергенции сенсорная информация передается
одновременно по многим параллельным каналам. Подобное устройство–видимо,
важнейшая из причин чрезвычайной надежности сенсорных систем. Они устойчивы к
потере отдельных нейронов в результате заболевания или старения организма.
Функции таких систем ухудшаются только при повреждении большого числа их
элементов.
Роль тормозных синапсов. В сенсорных путях не только присутствуют возбуждающие
синапсы, но используются и разные виды
торможения. В следующем разделе будет описана его роль при выделении
информации. Кроме того, торможение выполняет следующие функции.
1. Оно предупреждает неограниченное распространение возбуждения
по нейронной сети. Если торможение, осуществляемое глицинергическими синапсами,
блокировать антагонистом глицина стрихнином, нарушение упорядоченной передачи
информации в ЦНС приведет к конвульсиям и смерти.
2. Очень часто высшие сенсорные нейроны посылают
коллатерали к интернейронам, тормозящим сенсорные нейроны на более низком
уровне того же пути. Такое торможение по
типу обратной связи регулирует усиление при передаче информации. Вероятно,
такова же роль и «деполяризации первичных афферентов» в соматосенсорной
системе.
3. Наконец, высшие несенсорные мозговые центры могут
через нисходящие тормозные пути (нисходящее
торможение) блокировать передачу сигналов в сенсорных системах. Такие
механизмы позволяют, в частности, игнорировать некоторые элементы сенсорной
информации, когда внимание сфокусировано на других.
Неспецифичные системы. Не все нейроны, получающие информацию от сенсорных
систем, специфичны (т. е. связаны только с одной модальностью). Все сенсорные
системы связаны с мультимодальными, или
неспецифичными, нейронными группами и путями, к которым приходят сигналы от
нескольких из этих систем сразу. Важная неспецифичная система локализована в
зоне ретикулярных ядер ствола мозга и зрительного бугра (таламуса). По всей
вероятности, специфичные (унимодальные) сенсорные пути передают точную
информацию о стимулах (сообщают о том, что происходит), тогда как неспецифичные
ответственны за сенсорную интеграцию и определяемую стимулами модификацию
поведения (сообщают о значении происходящего). Зачастую его изменение
заключается в активации и перефокусировке внимания, относящихся к важным
функциям восходящей активирующей ретикулярной системы.

Рис. 8.7.
Простая модель сенсорной системы, иллюстрирующая принцип латерального
торможения. А. Две «молнии» символизируют
два одновременных стимула на малом расстоянии один от другого. Черным изображена анатомия сенсорной
системы. Числа соответствуют
относительному уровню возбуждения, в прямоугольниках
показаны серии потенциалов действия. Предполагается, что в каждом «прямом»
синапсе уровень возбуждения увеличивается втрое. В то же время коллатерали и
тормозные интернейроны обеспечивают латеральное торможение равное входной
величине без усиления. Результирующее возбуждение находят, вычитая уровень
торможения из уровня возбуждения (см. текст). Б. Распределение чистого возбуждения на трех этапах сенсорной
передачи
Выделение отличительных характеристик
сенсорной информации
Рецептивные поля первичных афферентных нейронов
определяются концевым ветвлением их аксонов, т.е. распределением и количеством
рецепторов, с которыми он соединен. Афферентное волокно возбуждается стимулами
в любом месте своего рецептивного поля. Однако, как правило, рецептивные поля центральных нейронов
устроены сложнее. Многие из этих нейронов возбуждаются стимулами в центре
рецептивного поля, но тормозятся стимулами, попадающими в его периферическую
зону, размеры и форма которой у разных нейронов неодинаковы. Это торможение –
результат соединений первичных афферентов с интернейронами, образующими тормозные
синапсы на вторичных нейронах. Поскольку торможение направлено «вбок» и
осуществляется нейронами того же сенсорного пути, где происходит и возбуждение,
оно называется латеральным
(боковым).
Сложные рецептивные поля центральных сенсорных
нейронов служат для выделения конкретных аспектов (признаков) общей сенсорной
информации. Одна из главных задач здесь
– усиление контраста. Ярче всего это проявляется в зрительной системе и
кожной механорецепции. Результат такого усиления контраста в том, что глаза
информируют нас не столько об абсолютных уровнях яркости, сколько о различиях в
ней, а следовательно, о границах между объектами.
Усиление контраста латеральным
торможением.
Схема сенсорной системы на рис. 8.7 сильно упрощена,
поскольку не отражает дивергенции и конвергенции, обсуждавшихся в предыдущем
разделе. «Молнии» изображают два точечных стимула, находящиеся на небольшом
расстоянии друг от друга и действующие на группу рецепторов одновременно.
Результирующее распределение возбуждения таково, что стимулы первоначально не
различаются как отдельные раздражители. Однако за счет подбора параметров
латерального торможения удается достичь их полного разделения через два этапа
синаптической передачи.
Чтобы смоделировать латеральное торможение, первичные
афференты на схеме на рис. 8.7 соединяются со следующими вторичными нейронами
не только непосредственно. Каждый из них возбуждает своими аксонными
коллатералями интернейроны (черные кружки на рисунке), а каждый из последних
тормозит соседний вторичный нейрон (белые кружки). Степень торможения,
естественно, зависит от степени возбуждения интернейронов, а значит, и
первичных афферентов. Модель предполагает, что каждому синапсу между первичным
и вторичным нейронами соответствует коэффициент усиления, равный 3; при этом
торможение посредством интернейронов (один возбуждающий и один тормозный
синапс, расположенные последовательно) дает общий коэффициент усиления — 1. Как
обычно в случае постсинаптического торможения, тормозные влияния (потенциалы)
вычитаются из возбуждающих. На следующем этапе синаптической передачи этот
процесс повторяется с теми же коэффициентами передачи. На рис. 8.7 уровни
возбуждения и торможения указаны цифрами. Возникающее в результате чистое
возбуждение можно понимать, например, как «число потенциалов действия» (в
красных рамках—гипотетические записи импульсации аксонов). Поэтапные изменения
легко вычислить с учетом приведенных выше допущений.
Выделение сложных признаков высшими
сенсорными нейронами. Усиление
контраста не единственный способ выделения информации высшими сенсорными
нейронами. В проекционных и ассоциативных областях коры головного мозга
отдельные нейроны выделяют из сенсорного возбуждения гораздо более сложную
информацию. Например, в соматосенсорной области есть клетки, кодирующие скорость
и направление движения раздражителя по коже. Зрительная кора содержит простые и
сложные клетки, выделяющие отдельные признаки геометрии и движения зрительных
стимулов. Более подробно организация различных сенсорных проекционных зон коры
обсуждается в главах, посвященных соответствующим сенсорным системам. Здесь,
говоря об общей сенсорной физиологии, важно подчеркнуть, что наши центральные
сенсорные системы, особенно корковые, анализируют входные сигналы, обеспечивая
их сознательное восприятие, путем
выделения, или абстрагирования, сенсорной информации.
8.3.
Общая субъективная сенсорная физиология
Основные характеристики ощущений
В начале этой главы субъективная сенсорная физиология
определялась как область исследований, занимающаяся соотношением между
физико–химическим миром стимулов и
субъективным миром ощущений и восприятии.
По традиции у ощущений различают четыре основных
характеристики –интенсивность,
качественный тип, временную длительность и пространственную протяженность. Для некоторых сенсорных
модальностей в последнем случае лучше говорить о местоположении. Если обратиться к слуху или обонянию, это означает
способность обнаружить (локализовать) источник звука или запаха. В случае
вкуса, например, пространственной характеристики вообще нет.
Прежде всего рассмотрим первые два из перечисленных
параметров. Начнем с качественного типа.
Дело в том, что ощущения принципиально различны по своей природе. «Видение» –
это нечто совершенно иное, нежели «слышание». Зрение, слух, обоняние, вкус и
т.д. называют сенсорными модальностями. Каждая из них включает различные качественные типы ощущений. Красный
цвет–один тип зрительных ощущений, зеленый–другой.
В классической медицине различали пять «чувств»:
зрение, слух, вкус, обоняние и осязание. Теперь выделяют несколько
дополнительных модальностей (например, чувства температуры и равновесия).
Вопрос о количестве присущих человеку «чувств» всегда будет связан с
особенностями интерпретации этого понятия.
Следует также иметь в виду, что некоторые виды
позвоночных обладают сенсорными органами, которых у нас нет. Например, лицевые
ямки некоторых змей содержат рецепторы инфракрасного излучения, позволяющие
обнаруживать теплокровную добычу. У некоторых рыб есть органы, чувствительные к
электрическим полям. Они улавливают токи, связанные с мышечной активностью
жертвы, которая зарылась в донный песок, и помогают ее локализовать.
Современная техника позволяет сконструировать аналогичные по свойствам приборы,
однако, чтобы мы могли ими пользоваться, информация на выходе должна быть
преобразована в зрительные или слуховые сигналы.
Все пять классических чувств человека относятся к
экстероцепции. О существовании проприоцепторов и интероцепторов не было
известно, пока физиологи их не открыли.
Особые трудности возникают при классификации боли и
других неприятных ощущений, например зуда. Боль–это сенсорная модальность, а
зуд, возможно, следует считать ее качественным типом. Однако она отличается от
других чувств тем, что присутствует во всех категориях ощущений–интеро–,
проприо– и экстероцепции. Ноцицепторы, воспринимающие боль, занимают среди
кожных рецепторов (все это экстероцепторы) особое положение, поскольку их
информация касается не внешнего мира, а повреждений собственного тела. Иными
словами, боль как модальность связана с телом. Подробнее физиология боли
рассмотрена в гл. 10.
Именно для качественной характеристики ощущений
Иоганнес Мюллер (1837) сформулировал закон
специфических сенсорных энергий, упоминавшийся выше: модальность зависит не
от стимула, а от раздражаемого им органа.
Сенсорные модальности нельзя сопоставлять друг с
другом с помощью прямых измерений. Однако на уровне качественных типов ощущений
такие измерения вполне возможны. Если медленно изменять частоту тона, будет
обнаружен некоторый порог, т.е. различие
в частотах, достаточное для заметного на слух перехода к другому качеству
звучания. Точно так же, меняя частоту электромагнитного излучения, можно
определить порог воспринимаемого изменения цвета.
Интенсивность ощущения: абсолютные и
дифференциальные пороги
Интенсивность ощущений наиболее подробно изучалась психофизическими методами. Интерес к
тому, можно ли измерить эту характеристику, привел Г. Фехнера к разработке
примерно в 1850 г. первой научной методики количественного описания
субъективного опыта. Он же вывел первый психофизический закон, устанавливающий
количественную связь между физической интенсивностью (φ) и силой ощущения (ψ). До этого господствовала
доктрина Декарта (1596–1650), отрицавшего возможность измерений в субъективной
сфере («res cogitans»).
Центральным понятием психофизики стало понятие сенсорного порога.
Абсолютный порог определяется как наименьший по интенсивности стимул,
способный вызвать определенное ощущение. Некоторые авторы обычно ограничивают
смысл этого термина, понимая под ним наиболее низкий порог, достижимый при
оптимальных условиях стимуляции и адаптации. Например, пороговые значения для
слуха зависят от частоты звука, а для зрения – от времени адаптации.
Дифференциальные пороги. В надпороговом диапазоне определяют еще один вид
порога–«едва заметное различие». Это величина, на которую один стимул должен
отличаться от другого, чтобы их разница воспринималась человеком. Э. Вебер
(1834), экспериментируя с ощущением силы, первым показал, что для различения
веса двух предметов их разница должна быть больше, если оба они тяжелые, и
меньше, когда оба легкие. На рис. 8.8,A показано
соотношение между предъявляемым весом и его едва заметным увеличением. В
среднем диапазоне такой стимуляции зависимость линейная;
иными словами, чтобы превысить дифференциальный порог,
требуется одинаковая часть начального веса. Минимальное различимое изменение
интенсивности стимуляции (Δφ) составляет постоянную долю (с) ее
исходной интенсивности (φ). Это закон Вебера, выражаемый уравнением
Δφ
/ φ = с; Δφ = с φ. (1)
Значит, Δφ / φ –константа для всех
интенсивностей стимула. Данное правило, действительно, выполняется в широком их
диапазоне для многих сенсорных модальностей. Однако по мере приближения к
абсолютному порогу коэффициент Вебера
(Δφ / φ) обычно растет. Это проиллюстрировано на рис. 8.8,Б для
громкости тона. Очевидно, в этом случае закон Вебера начинает действовать
только при интенсивности стимула на 40 дБ выше абсолютного порога: начиная с
этого уровня, коэффициент Вебера остается постоянным. Аналогичные кривые
получены и для других модальностей.
Коэффициент Вебера – полезная мера относительной
чувствительности сенсорных систем. Нельзя математически сравнить
чувствительность глаза к силе света с чувствительностью уха к уровню звукового
давления, но можно сопоставить между собой безразмерные коэффициенты Вебера для
этих модальностей. В подобных опытах глаз несколько лучше, чем ухо, улавливает
разницу адекватных стимулов по интенсивности.
Для соответствия экспериментальным данным закон Вебера
можно переформулировать следующим образом:
Δφ
/ (φ+a) =c
Δφ=
c (φ+a)
(2)
где а – константа, обычно небольшая по численному
значению. Когда интенсивность стимула (φ) достаточно велика, членом «а»
можно пренебречь, и выражение, соответствующее исходной форме закона Вебера,
описывает наблюдения достаточно точно (рис. 8.8, В).
В некотором смысле константа «а»–просто поправочный
член, обеспечивающий более точное соответствие экспериментальным данным. Однако
его можно рассматривать и как «шум» в сенсорном канале. С точки зрения
нейрофизиологии такой «шум» объясняется спонтанной активностью сенсорных
нейронов, особенно высших уровней, т. е. их импульсацией в отсутствие
действующих на сенсорный орган стимулов. Вероятно, она добавляется к активности
нейронов в ответ на стимуляцию, поэтому также влияет на порог, необходимый для
едва заметного увеличения сигнала в ЦНС. Спонтанная активность мала по
сравнению с реакцией на сильные стимулы, однако, когда стимул слаб, влияет на
величину (Δφ / φ). В соответствии с этой нейрофизиологической
интерпретацией, абсолютный порог должен быть равен минимальной интенсивности
стимула, достаточной для возбуждения сенсорной системы,

Рис. 8.8.
Коэффициент и закон Вебера. А.
Соотношение между величиной раздражения (φ) и его
увеличением (Δφ), необходимым для того, чтобы превзойти
дифференциальный порог ощущения силы. 6. Зависимость коэффициента Вебера
(Δφ/φ) от интенсивности звуковой стимуляции. Коэффициент Вебера
становится постоянным только при громкости более чем на 40 дБ выше абсолютного
порога. В. Коррекция коэффициента
Вебера с помощью константы «а». Исправленный закон применим и к околопороговым
стимулам (А. Б: по [1] с изменениями)
различимо большего спонтанной активности. Данный
подход используется в некоторых психофизических теориях (например, в
рассматриваемой ниже «сенсорной теории принятия решений»). В разделе,
посвященном интегративной сенсорной физиологии, мы вернемся к вопросу о том,
действительно ли спонтанная активность решающим образом влияет на абсолютный
порог.
Помимо определения порогов, можно ли как–то еще
измерить субъективные ощущения? Фехнер предложил использовать дифференциальные
пороги

Рис. 8.9.
Схематическое изображение закона Фехнера. По оси абсцисс–интенсивность стимула, по оси oрдинат–
дифференциальные пороги
для построения
шкалы интенсивности ощущений (ψ). Нуль на ней соответствует абсолютному порогу, следующее ощущение
больше точно на величину одного едва заметного различия, следующее – еще на одну
такую величину и т.д. Поскольку каждая ступенька соответствует наименьшему
возможному усилению ощущения, этот прирост и берется за основную единицу
интенсивности ощущения. На рис. 8.9 показан график зависимости интенсивности
ощущения от интенсивности стимула. Это соотношение между ψ
и φ описывается уравнением, известным как психофизический закон Фехнера. Поскольку стимул увеличивается
путем последовательного прибавления все возрастающей величины Δφ,
получается логарифмическая кривая (рис. 8.9). Отсюда закон утверждает: при
линейном увеличении интенсивности ощущения (ψ) интенсивность стимула
(φ) растет логарифмически. Если психофизическую функцию (рис. 8.9)
представить в линейно–логарифмической системе координат, кривая превратится в
прямую (рис. 8.13). Фехнеровский закон выражается уравнением
ψ = k • log (φ / φо), (3)
где ψ –интенсивность ощущения, k–константа,
φ – интенсивность стимула и (φо–его абсолютный порог. Данный
психофизический закон основан на двух главных допущениях.
1. Он действует при справедливости закона Вебера. При очень
слабой стимуляции веберовский коэффициент не постоянен. Значит, закон Фехнера в
этой области не действует.
2. Допускается также, что все приросты интенсивности
ощущения равны, т. е. что пороговое изменение слабого и сильного стимулов
ощущается одинаково. Рассматривая закон Стивенса, мы убедимся в некорректности
данного допущения; в результате справедливость «закона» Фехнера сильно
ограничена. В свете этих данных лучше было бы говорить о психофизическом соотношении Фехнера.
Заслуга Фехнера в том, что он первым ввел пригодную
для использования шкалу «интенсивности ощущения». Он вполне заслужил титул отца
экспериментальной психологии. Теперь мы знаем, что ордината Ψ в законе Фехнера выражает скорее
разрешающую способность, чем интенсивность ощущения. Ограничения этой шкалы
обсуждаются ниже.
Соотношение Фехнера использовалось для выведения
логарифмических систем мер, применимых в сенсорной физиологии, например шкалы децибелов и фонов. В промежуточном
диапазоне частот и интенсивностей звуков минимальное заметное различие
соответствует примерно 1 дБ.
В предыдущем разделе было показано, что абсолютные и
дифференциальные пороги относятся к фундаментальным психофизическим понятиям.
Теперь вернемся к вопросу о том, как их измерять. Примеры относятся к
абсолютным порогам, но применимы и к дифференциальным.
Реакции биологических систем неустойчивы, поэтому при
тестировании слабыми стимулами испытуемый иногда их воспринимает, иногда–нет.
Следовательно, нельзя определить порог как интенсивность стимула, ниже которой
он никогда не ощущается, а выше–всегда заметен. Стимул нужно предъявлять
несколько раз, после чего, обработав результаты статистически, оценить
«истинный», средний порог. Есть несколько методик определения порога, некоторые
из которых разработаны еще Фехнером.
К классическим относится метод пределов. Исходный стимул здесь далек от предполагаемого
порогового значения. Например, интенсивность сначала устанавливают такой
высокой, что испытуемый легко воспринимает стимул, а затем снижают до тех пор,
пока она не окажется подпороговой. Затем тестирование начинают снова–на этот
раз с очень слабых стимулов, которые усиливают до достижения порога. Такие
восходящие и нисходящие последовательности повторяют несколько раз и берут
среднее значение полученных пороговых величин. Интенсивность стимула можно
записывать автоматически на миллиметровой бумаге, так что для выявления
колебаний порога во времени необходимо будет лишь провести линию, соединяющую
концы отрезков получаемых кривых.
Субъективная сенсорная физиология и
поведенческие эксперименты на животных.
Метод пределов, как и другие способы измерения порогов, можно использовать и в
экспериментах на животных.

Рис. 8.10. Измерение
зрительного порога голубя в поведенческом эксперименте. А. Схема экспериментальной установки. Голубь клюет ключ А, если
видит свет, в результате чего сила света уменьшается. Клевание ключа Б, когда
света не видно, увеличивает интенсивность последующего светового стимула. Б. Постепенное снижение порога после
выключения яркого фонового освещения. Получена кривая темповой адаптации голубя (по [8, 9] с изменениями)
Пример показан на рис. 8.10. У голубя выработан
условный рефлекс (см. гл. 6) получать пищевое вознаграждение после клевания
ключа А, если он видит свет, и ключа Б,
если света не замечает. При этом ключ А снижает интенсивность светового
стимула, а ключ Б увеличивает ее. В ходе такого поведенческого эксперимента
получаются восходящие и нисходящие кривые силы света, точки перегиба на которых
соответствуют зрительному порогу. Если клетку с голубем изолировать от света,
то непрерывное измерение порога таким способом даст кривую темновой адаптации,
сходную с получаемой для человека. Этот пример ясно показывает, что методы
субъективной сенсорной физиологии применимы и в поведенческих экспериментах на
животных.
Подобные эксперименты упоминаются здесь в разделе о
субъективной сенсорной физиологии, поскольку своими методами они очень сходны с
психофизическими опытами на людях. Можно, естественно, считать, что измеренные
параметры поведения отражают субъективные ощущения животных, которые вовсе не
обязательно сравнимы с сознательными переживаниями человека.
В одной из модификаций метода пределов испытуемый сам
управляет стимуляцией (например, поворачивая рукоятку), усиливая ее до тех пор,
пока она не станет ощущаться, или ослабляя до невоспринимаемого уровня.
Преимущество данного метода настройки
в более активной роли испытуемого по сравнению с другими психофизическими
процедурами, при которых неизбежна усталость, вызываемая монотонностью
эксперимента. Недостаток его–в невозможности выдержать одно и то же время
стимуляции.
Психометрическая функция. Гораздо сложнее процедура предъявления различных по
интенсивности стимулов в случайном порядке. Однако она позволяет получить более
точные данные. Это так называемый метод
постоянных стимулов. Они подбираются таким образом, что самый слабый почти
никогда не замечается, а самый сильный ощущается почти всегда. После каждого
предъявления стимула испытуемый отвечает, почувствовал он его или нет, и
регистрируется процент обнаруженных стимулов каждой интенсивности. На рис. 8.11
показаны примеры полученных таким способом результатов. Точки соответствуют
относительным частотам выявления стимулов разных интенсивностей. У большинства
испытуемых проходящая через них кривая
психометрическая функция–S–образная. Порогом считается величина стимула, при
которой он обнаруживается в 50% случаев. На рис. 8.11 порог не совпадает ни с одним
из предъявленных стимулов и определяется путем интерполяции.
S–образная психометрическая функция часто по форме
совпадает с кумулятивной кривой нормального распределения (интегралом гауссова
распределения). Если относительные частоты обнаружения случайно предъявляемых
стимулов наносятся на график как вероятности отклонения от среднего (величина z по правой оси ординат), экспериментальные точки
образуют примерно прямую линию (рис. 8.11,Б). To, что психометрическая функция зачастую описывается
S–образной кривой, вызывает определенный теоретический интерес. Значит, в
основе флуктуации в восприятии лежит статистический процесс (см. также
следующий раздел).
Статистический анализ порогов. Сенсорная
теория принятия решений
В предыдущем разделе мы гипотетически определили
абсолютный порог как интенсивность
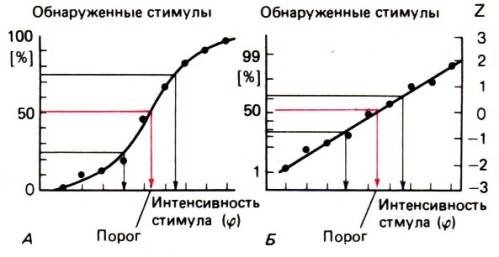
Рис. 8.11.
Психометрическая функция, получаемая при измерении пороговой интенсивности
посредством случайного предъявления стимулов (метод «постоянных стимулов»).
Порог определяется как точка на
кривой, соответствующая обнаружению стимула в 50% случаев. А. Зависимость относительной частоты успешных попыток различения
(ось ординат) от интенсивности
стимуляции (ось абсцисс). Б. Зачастую
S–образная психометрическая функция соответствует интегралу нормального
(гауссова) распределения. Если относительную частоту успешных попыток
преобразовать в величины Z (отклонение от среднего), психометрическая функция
принимает вид прямой линии (по [1] с изменениями)
стимула, приводящую к возбуждению сенсорного канала,
едва заметно превышающему уровень спонтанной активности его нейронов. Если
предположить, что флуктуации этой активности и изменчивость ответов на слабый
(постоянный и повторяющийся) стимул описываются кривой нормального
распределения, то получается частотный график типа показанного на рис. 8.12, А.
Из него ясно, что наблюдатель, у которого возникает нейронное возбуждение на
уровне «а», не может с уверенностью судить, соответствует ли оно спонтанной
активности или же воздействию слабого стимула. В случае сильных стимулов такой
проблемы нет, поскольку распределение ответов на них не перекрывается со
спонтанной активностью.
Проблемы
идентификации околопороговых
стимулов на фоне шума рассматриваются «сенсорной теорией принятия решений»
(теорией обнаружения сигналов) [15, 21]. В соответствии с ней, наблюдатель,
пытающийся выделить слабый сигнал из шума, должен принять решение. Оно может,
например, состоять в том, что все уровни нейронного возбуждения, превышающие
«б», нужно считать стимулом, а все более низкие–спонтанной активностью.
Разделительная линия, соответствующая такому решению, называется критерием или
отклонением. В примере, иллюстрируемом рис. 8.12, А, решения, исключающие
ошибку, невозможны. В случаях, соответствующих равномерно закрашенным (красным
цветом) зонам, стимулы не обнаруживаются, а в заштрихованной области справа от
критерия спонтанная активность ошибочно воспринимается как ответ на стимул.

Рис. 8.12.
Схема предъявления околопорогового стимула, иллюстрирующая сенсорную теорию принятия
решений. А. Распределения спонтанной
активности («шума») в сенсорной системе и нейронных ответов на многократное
предъявление постоянного околопорогового стимула. Б. Возможное сочетание правильных и неправильных ответов.
Подробности в тексте
На рис. 8.12,Б показаны все четыре возможные
комбинации. В данном случае два правильных ответа: «хватай» и «правильный
отказ». Есть и два неправильных сочетания: «ложная тревога» и «упустил».
Вероятности этих ответов на слабые стимулы зависят друг от друга. Для двух
правильных они связаны обратной пропорциональной зависимостью–чем больше
правильных «хватаний», тем меньше правильных отказов. То же самое справедливо и
для неверных ответов–чем больше «упусканий», тем меньше ложных тревог. Это
легко заметить, если на рис. 8.12,А сдвинуть критерий: площади двух
заштрихованных зон изменяются обратно пропорционально.
Сенсорная теория принятия решений по–своему определяет
порог. Из нее следует, что он зависит от двух
факторов–сенсорной избирательности и критерия, установленного в процессе принятия решений.
Величина избирательности наиболее близка к пороговому значению. Методы
исследования избирательности сенсорной системы и процессов принятия решений (у
человека и животных) описаны в работах [15, 21].
Смысл того, что при неопределенной стимуляции
сенсорные ответы зависят от критериев принятия решений, выходит далеко за рамки
сенсорной физиологии.
При более общем подходе шум определяется как сумма
факторов неопределенности. В качестве примера возьмем медицинскую диагностику.
Неопределенное зрительное впечатление о том, что у кожи больного желтоватый
оттенок, поставит врача перед дилеммой–желтуха это или нет; все зависит от
степени желтизны, т.е. от критерия принятия решения. Он, разумеется, зависит от
опыта доктора–от его интуитивной оценки вероятности. Однако решение зависит и
от оценки его последствий. Если предполагаемое заболевание серьезно, а проверка
диагноза не слишком затруднительна, врач предпочитает скорее дать «ложную
тревогу», нежели «упустить». Если материальные и нематериальные затраты,
связанные с решением, распределены по–другому, соответствующим образом
сместится и критерий. Иными словами, принятие решений в медицине обусловлено не
только анализом вероятностей, но и оценкой последствий, а также этическими
принципами.
В течение примерно ста лет серьезных попыток оспорить
допущение Фехнера относительно измерения интенсивности ощущений в единицах
дифференциального порога не было. Однако Стивенс показал, что этот порог не
постоянен. Например, тон, громкость которого на 20 порогов выше абсолютного,
звучит для нас более чем вдвое громче тона той же частоты с громкостью на 10
порогов выше абсолютного. Значит, едва заметное различие в интенсивности не
одинаково для всего ее диапазона. Стивенс пришел к заключению, что
психофизический закон требует шкалы ощущений другого вида. В его попытках
измерить их непосредственно понятие дифференциального порога уже не занимает
значительного места.
Прежде чем мы перейдем к рассмотрению психофизического
закона Стивенса, следует описать другой случай, который кажется впечатляющим
аргументом в пользу позиции Фехнера. В течение более 2000 лет астрономы
наблюдают и классифицируют звезды. Приблизительно в 150 г. до н. э. греческий
астроном Гиппарх ввел количественную шкалу их яркости, которая используется и
сегодня. Самые яркие звезды принадлежат к первому классу, следующие по
яркости–ко второму и т.д. вплоть до звезд шестого класса, которые уже едва
различимы невооруженным глазом. В течение многих столетий астрономы
пользовались этой шкалой, пока не появились фотометрические приборы. Таким
образом, классификацию звезд можно считать широкомасштабным психофизическим
экспериментом продолжительностью в сотни лет. К концу XIX в., когда ученые
сопоставили эту шкалу с данными измерений яркости, они обнаружили
приблизительно логарифмическую зависимость, соответствующую закону Фехнера. Она
представлена на рис. 8.13. Сейчас звезды классифицируют в соответствии с их
фотометрической яркостью, однако логарифмическая шкала звездных величин все еще
широко используется.
Закон Фехнера применим в этом случае потому, что
астрономы не оценивали интенсивность
своих ощущений, а ориентировались на критерий различимости. Главным при такой классификации было то, что звезда
первой величины заметно ярче, чем второй, и т.д. Насколько велика разница в
ощущениях, не учитывалось.

Рис. 8.13.
Средняя яркость звезд разной величины при использовании линейной (А) и логарифмической (Б) шкал по оси абсцисс (данные Jastron из [24])
Значит, можно сказать, что закон Фехнера применим в
том случае, когда используется субъективная шкала
различимости. По природе она чисто порядковая–с ее помощью невозможно перемножать
.(например, утверждать, что нечто «вдвое» ярче). С другой стороны, шкала
Стивенса описывает именно отношения
между величинами. Это необходимо для статистических операций, например
вычисления арифметического среднего и стандартного отклонения. В табл. 8.1
приводятся математические действия, допускаемые различными шкалами. Они
перечислены в восходящем порядке, так что статистические операции, возможные на
более низком уровне, выполнимы и на более высоком, но не наоборот.
В психофизических экспериментах по Стивенсу
испытуемого просят оценить интенсивность своих ощущений непосредственно. Она не
вычисляется путем подсчета количества едва заметных различий. Разработаны
различные методы подобных прямых
психофизических оценок. Согласно одному из них, интенсивность оценивается
численно: за нуль берется абсолютный порог, а другие цифры описывают ощущение,
вызываемое стандартным стимулом. Важно, что данная шкала не разбита на отрезки,
а допускает постепенные (градуальные) изменения. Испытуемый использует такие
понятия, как «наполовину» или «вдвое» интенсивнее, и отмечает это
соответствующими числами. Данный метод основан на принципе пропорциональности.
Полученные результаты заставили Стивенса предположить, что соотношение между
интенсивностями ощущения (Ψ) и стимула (φ ) описывается степенной функцией. В соответствии с
законом Стивенса
Ψ = k
(φ-φ0) (4)
где ψ–интенсивность ощущения, к–
константа, зависящая от шкалы, φ– интенсивность стимула, a φ0–его интенсивность при абсолютном пороге. Показатель
степени а, зависящий от сенсорной модальности и условий стимуляции, определяет
форму кривой зависимости ψ от φ. Например, если он равен 1, соотношение линейное.
Если он больше, сила ощущения возрастает быстрее интенсивности стимула (кривая
загибается вверх), а если меньше, то наоборот.
Вычертив, соответствующие графики в логарифмической
системе координат, получим прямые линии
logΨ = logk + a–log(φ-φ0) (5)
Из уравнения (5) следует, что данные попадают на
прямую линию с координатами log(φ-φ0) и logΨ. Ее
наклон (тангенс угла с осью абсцисс) соответствует показателю степени а.
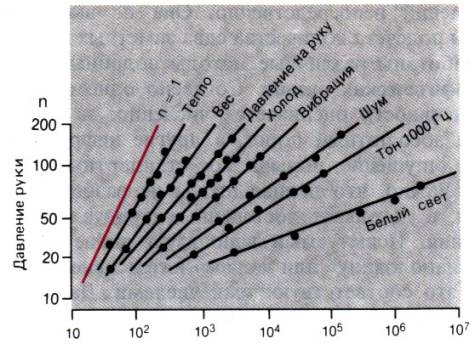
Степенные функции, выведенные Стивенсом для разных
модальностей и качественных типов ощущений, показаны на рис. 8.14.
Соответствующие коэффициенты, несомненно, зависят от условий
Таблица 8.1. Виды шкал и операций, которые они допускают (по [24]
с изменениями)